
























































































General Neurology
Neurologic manifestations of celiac disease and gluten sensitivity
Jan. 23, 2025
MedLink®, LLC
3525 Del Mar Heights Rd, Ste 304
San Diego, CA 92130-2122
Toll Free (U.S. + Canada): 800-452-2400
US Number: +1-619-640-4660
Support: service@medlink.com
Editor: editor@medlink.com
ISSN: 2831-9125
Toll Free (U.S. + Canada): 800-452-2400
US Number: +1-619-640-4660
Support: service@medlink.com
Editor: editor@medlink.com
ISSN: 2831-9125
Worddefinition
At vero eos et accusamus et iusto odio dignissimos ducimus qui blanditiis praesentium voluptatum deleniti atque corrupti quos dolores et quas.
Patients and clinicians frequently overlook disorders of olfaction, but they can worsen quality of life, distort taste, lessen appetite, augment depressive disorders and sometimes paranoia, and can also pose a safety risk. When patients report alterations in the quality of olfaction in response to an odorant (ie, parosmias), the perceptions are almost universally unpleasant, a condition referred to as aliosmia (the perception of unpleasant odors from nominally pleasant odorants). Disorders of olfaction are particularly common with synucleinopathies, aging, and COVID-19. This article reviews the range of olfactory symptoms, categorizes disorders of olfaction, prognosis, and complications of these disorders, and reviews disease pathogenesis, diagnosis, and management.
|
• Patients with olfactory symptoms generally report diminution or absence of olfactory sensation (hyposmia or anosmia, respectively) and forms of distorted olfaction (parosmia). | |
|
• Except in unusual circumstances, hyperosmia is a subjective sensation of hyperacuteness of olfaction. | |
|
• There is no evidence that pregnant women or migraineurs experience an objective increase in olfactory sensitivity. | |
|
• When patients report alterations in the quality of olfaction in response to an odorant (ie, parosmias), the perceptions are almost universally unpleasant, a condition referred to as aliosmia (the perception of unpleasant odors from nominally pleasant odorants). | |
|
• Aliosmias may involve the perception of fecal or rotten smells (cacosmia) or chemical or burned smells (torquosmia). | |
|
• Complaints of impaired “taste” are often a symptom of olfactory dysfunction because much of the flavor of a meal derives from olfactory stimulation. Indeed, the complex sensory experience of “flavor” during the consumption of foods and drinks cannot be constructed simply from combinations of the basic taste qualities (sweet, salty, sour, bitter, and umami/savory). | |
|
• Chemosensory deficit may be the first symptom (a "sentinel symptom") in patients with COVID-19, but there is wide variation in the proportion of cases in which this is reported to occur. | |
|
• Most patients with COVID-19-related chemosensory dysfunction do not present associated nasal congestion or rhinorrhea. | |
|
• Presbyosmia (literally “elderly olfaction” or “old age olfaction”) is the gradual loss of olfactory abilities that occurs in most people as they grow older. | |
|
• Clinically significant olfactory loss is common in the elderly but frequently unrecognized, partly because deficits typically accumulate gradually over decades. Indeed, self-reported olfactory impairment significantly underestimates prevalence rates obtained by olfactory testing. | |
|
• Olfactory deficits involving odor detection, identification, and discrimination are present in more than 90% of patients with early-stage Parkinson disease. | |
|
• In dementia with Lewy bodies, as in Parkinson disease, olfactory dysfunction is nearly universal, develops early (before any movement or cognitive disorder), and is often severe. |
Printed medical illustrations began in 1490, and by the beginning of the 16th century, they included representations of afferent connections from the special sensory organs to the brain (129). These were typically part of highly schematic diagrams of brain function representing the medieval cell doctrine. Three “cells” or ventricles were usually assigned functions of sensory integration and imagination, cognition, and memory (130). Indeed, many early 16th-century woodcuts of the medieval cell doctrine show presumptive connections between the organs subserving the special senses, either with the most anterior cell or ventricle of the brain or with a specific portion of it--the sensus communis (ie, sensory commune or common sense, a structure Aristotle had postulated is responsible for monitoring and integrating the panoply of sensations from which unified conscious experience arises) (129; 130; 131). A representation of the olfactory bulbs is incorporated into many of these woodcuts, beginning with an illustration by German physician, philosopher, and theologian Magnus Hundt (Parthenopolitanus, 1449-1519) in 1501 in his Antropologium, which showed central projections of the two olfactory bulbs joining in the meshwork of the rete mirabile (131; 132). German physician and anatomist Johann Eichmann, known as Johannes Dryander (1500-1560), modified Hundt’s figure for his own monograph in 1537 but retained the representation of the olfactory bulbs (131; 132).
Illustration by German physician, philosopher, and theologian Magnus Hundt (1449-1519) in 1501 in his Antropologium. (Source: Hundt M. Antropologium de ho[min]is dignitate, natura, et p[rop]rietatibus, de elementis, pa...
German Carthusian humanist writer Gregor Reisch (c 1467-1525) published an influential and highly copied woodcut in his Margarita philosophica (1503), showing connections from the olfactory bulbs overlying the bridge o...
In 1503, German Carthusian humanist writer Gregor Reisch (c 1467-1525) published an influential and highly copied woodcut in his Margarita philosophica, showing connections from the olfactory bulbs overlying the bridge of the nose to the sensus communis in the anterior cell or ventricle (131). In the following centuries, numerous authors derived similar figures from Reisch’s original schematic illustration of the medieval cell doctrine, including Brunschwig (1512, 1525), Głogowczyk (1514), Romberch/Host (1520), Leporeus/Le Lièvre (1520, 1523), and several others (131).
Representations of the medieval cell doctrine from Polish philosopher and polymath Jan Głogowczyk (c. 1445?-1507), published posthumously in 1514. The 1514 image is derived from Reisch’s (1503) image and shows the Reisch scheme...
Representation of the medieval cell doctrine (1520) from German Dominican Johannes Romberch (Johann Horst von Romberch; c. 1480-1532/1533). Note the engraved connections from the special sense organs to the brain, particularly ...
Woodcut of the medieval cell doctrine from French author Guillaume Le Lièvre (Gulielmus Leporeus; fl. c. 1520). The olfactory bulbs are depicted as a pair of round structures at the bridge of the nose, and both show projection...
Similar representations were provided by Peyligk (1518) and Eck (1520) (131).
Copperplate engraving by Peter Weinher the Elder (1572). (Source: Staatliche Graphische Sammlung, München, via Wikimedia Commons. See: Lanska DJ. Representations of the olfactory bulb and tracts in images of the medieval cell d...
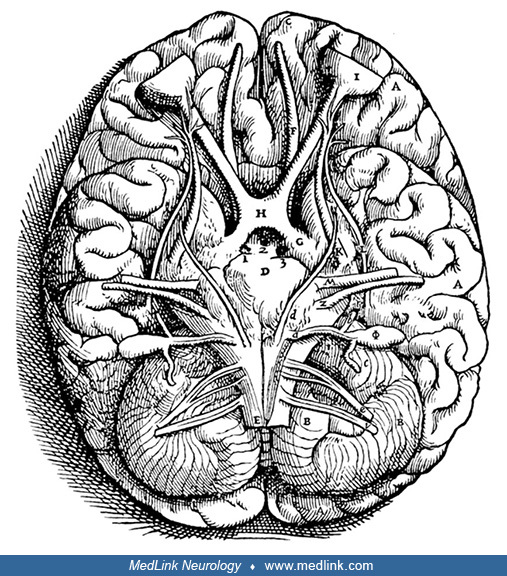
This is a unique representation of the medieval cell doctrine and the special senses because it not only shows the connections of some of the special sense organs to the brain (nose, eyes, and ears but not tongue) but also the ...
These stereotyped 16th-century schematic images typically located the olfactory receptors (depicted as small circular or oval objects resembling tiny eyeglasses) across the bridge of the nose and at exactly the anatomic level of the olfactory bulbs. Such images linked the olfactory bulbs to olfaction before the advent of more realistic images beginning in the mid-16th century.
German physician and anatomist Johann Eichmann, known as Johannes Dryander (1500-1560), modified Hundt’s figure for his own monograph in 1537 but retained the representation of the olfactory bulbs. (Source: Dryandrum J. Anatomi...
Observational anatomy was largely lost from the time of Galen in the second century, and it became regimented and dogmatized with the scholasticism of the Middle Ages until a few anatomists began to seriously challenge Galen beginning in the 16th century. Most notably, Flemish anatomist Andreas Vesalius (1514-1564) provided much greater realism with the publication of his de Humani corporis fabrica (1543); however, Vesalius’ image did not, in fact, show clear bulb-like enlargements but rather an optic tract and bulb of roughly uniform thickness.
(Source: Vesalius' de Humani corporis fabrica. [1543]. Courtesy of the U.S. National Library of Medicine, Bethesda, Maryland. Woodcut print edited by Dr. Douglas J Lanska.)
Even after Vesalius championed a return to observational anatomy, the medieval cell doctrine and its associated representation of the olfactory pathways persisted well into the 19th century, even if it gradually moved to the fringes of medical thought (131). These included, for example, Venetian humanist Lodovico Dolce's (1508/1510-1568) and Basque Franciscan Bernardus de Lavinheta's (died c 1530) edition of a much earlier work by Bernardus de Lavinheta Ramón Lull (Raimundus Lullius, c 1235-1316) in 1612 (131). One of the last of these representations was published in 1835 by British physician and phrenologist John Elliotson MD FRS (1791-1868) (131).
Representation of the medieval cell doctrine (1562) from Venetian humanist writer Lodovico (or Luigi) Dolce (1508/10-1568). This is a close copy of the representation of the medieval cell doctrine by Romberch (1520). Note the e...
From the Kingdom of Majorca on the east coast of Spain. Copperplate engraving, sixteenth or seventeenth century. (Source: Dibner Library of the History of Science and Technology, Smithsonian Institution, Washington, D.C. See: L...
Persistent early 17th-century representation of the medieval cell doctrine in Basque Franciscan Bernardus de Lavinheta's (died c. 1530) edition of a much earlier work by Bernardus de Lavinheta Ramón Lull (Raimundus Lullius; c....
Many of the histological features of the olfactory epithelium, the olfactory sensory nerves, the passage of olfactory nerves from the olfactory epithelium through the cribriform plate, the synapse of these bipolar neurons in the glomeruli of the olfactory bulb, and further circuits within the olfactory bulb were elaborated remarkably well in the late 19th century and early 20th century by Italian histologist Camilo Golgi (1843-1926) and his upstart nemesis, the Spanish histologist Santiago Ramón y Cajal (1852-1934) along with Cajal's disciples Tomás Blanes Viale (1878-1900) and Fernando de Castro (1896-1918) (85; 184; 185; 186; 187; 14; 169; 139; 48; 49; 50; 51; 151; 127; 133; 147; 214; 47; 76; 166; 197; 66).
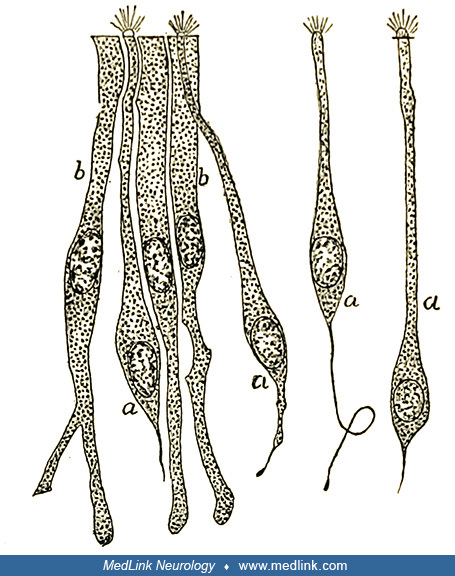
Santiago Ramón y Cajal's drawing of an anteroposterior section of the olfactory bulb and nasal mucosa of a newborn rat. Legend: (A) Olfactory epithelium situated below the cribriform plate; (a) bipolar cell, (b) epithelial or s...
Fernando de Castro (1896-1918) drawing of human olfactory bulb stained with Cajal’s gold chloride sublimate method. Legend: (A) Superficial substratum of the molecular layer with numerous cephalopodic cells. (B) Deep substratum...
Fernando de Castro (1896-1918) drawing illustrating the glomerular layer of the adult dog stained by the Cajal’s gold chloride sublimate method. Legend: (g) Intraglomerular fibrous elements. (a) Radioglomerular corpuscles. (f) ...
The greatness of these early histologists can be appreciated by comparing the drawings of histological preparations from the late 19th century with modern photomicrographs of histological and immunohistological preparations.
Olfactory epithelium labeled E18 cells after in-utero electroporation of an EGP-expressing plasmid injected into the olfactory placode at E14 (green). Hoechst (blue). Sagittal mouse section. a-e, n correspond to the counterpart...
Immunohistochemical staining for Dab1 protein (reelin signaling mediator, green) and Map2a,b protein (microtubule-associated protein, red) on mouse olfactory bulb sections at P3. Nuclei counterstained with Hoechst (blue). Legen...
Olfactory sensory neurons (green) at E17, labeled after in-utero electroporation of an EGP-expressing plasmid into the olfactory placode at E11. Nuclei counterstained with Hoechst (blue). Sagittal mouse brain section. ...
Axonal arborizations of olfactory sensory neuron axons in several glomeruli stained with the Golgi method. (Source: Figueres-Oñate M, Gutiérrez Y, López-Mascaraque L. Unraveling Cajal's view of the olfactory system. Front Neuro...
Various expressions are used by patients to describe disorders of smell or olfaction (Table 1).
|
Category |
Term | ||
|
Disturbance of perception (any) |
Dysosmia | ||
|
Alteration of quantity | |||
|
Absent |
Anosmia | ||
|
Decreased |
Hyposmia | ||
|
Normal |
Normosmia | ||
|
Alteration of quality | |||
|
Distorted (any) [general term] |
Parosmia | ||
|
Specific distortions | |||
|
1. Any unpleasant distortion of smell |
Aliosmia | ||
|
Hallucination2 |
Phantosmia | ||
|
1. "Unusual and unexpected but not necessarily foul or obnoxious" (93) | |||
Patients with olfactory symptoms generally report diminution or absence of olfactory sensation (hyposmia or anosmia, respectively) and forms of distorted olfaction (parosmia). Except in unusual circumstances, hyperosmia is a subjective sensation of hyperacuteness of olfaction. There is, for example, no evidence that pregnant women or migraineurs experience an objective increase in olfactory sensitivity.
When patients report alterations in the quality of olfaction in response to an odorant (ie, parosmias), the perceptions are almost universally unpleasant, a condition referred to as aliosmia (the perception of unpleasant odors from nominally pleasant odorants). Aliosmias, for example, may involve the perception of fecal or rotten smells (cacosmia) or of chemical or burned smells (torquosmia).
The terminology for alterations in the quality of olfactory perception (eg, cacosmia, torquosmia) can also be employed with hallucinated odors (phantosmias). Thus, one can speak of a "cacosmic phantosmia" for the hallucination of a fecal odor or a "torquosmic phantosmia" for the hallucination of a burned odor (94).
In addition, complaints of impaired “taste” are often symptoms of olfactory dysfunction because much of the flavor of a meal derives from olfactory stimulation. Indeed, the complex sensory experience of “flavor” while consuming foods and drinks cannot be constructed simply from combinations of the basic taste qualities (sweet, salty, sour, bitter, and umami/savory).
While chairman of the Department of Ophthalmology at Harvard Medical School, American ophthalmologist David G Cogan (1908-1993) distinguished "irritative" and "release" hallucinations in patients without psychosis in an influential paper in 1973 (43).
An "irritative" mechanism for hallucinations is supported by any of the following: (1) stereotyped content; (2) lack of awareness of the hallucinatory nature of the perception ("hallucinosis"); and (3) evidence of an irritative process (eg, migraine, tumor, seizures) (43; 25). In general, a "release" mechanism for hallucinations is supported by (1) a sensory deficit in the same modality as unimodal hallucinations, with onset of hallucinations in conjunction with or following the sensory deficit; (2) variable content; (3) awareness of the hallucinatory nature of the perception; and (4) no evidence of seizures or other irritative phenomena (including no other positive motor or sensory phenomena, not paroxysmal in character, and no epileptiform discharges on electroencephalography) (43; 135; 25). Release hallucinations can occur in normal individuals with pansensory deprivation (96; 95). Similarly, modality-specific release hallucinations can occur experimentally (95; 157) or pathologically (92; 135; 134) with unimodal sensory deprivation.
Congenital anosmia. Individuals with congenital anosmia typically lack any olfactory epithelium. In a case series of congenital anosmia involving detailed chemosensory evaluation followed by the performance of biopsies of the olfactory region, olfactory epithelium was not found in any of the biopsy specimens (112), suggesting that either the olfactory placode does not form normally or that it degenerates and is replaced with respiratory epithelium. Two olfactory placodes arise as thickened ectoderm from the frontonasal process. In the sixth week of development, the center of each placode grows inwards to form the two nasal pits. The invaginations give rise to the olfactory epithelium that lines the roof of the nasal cavity, whereas the raised margins are divided into medial and lateral nasal processes that give rise to the nose, the philtrum of the upper lip, and the primary palate.
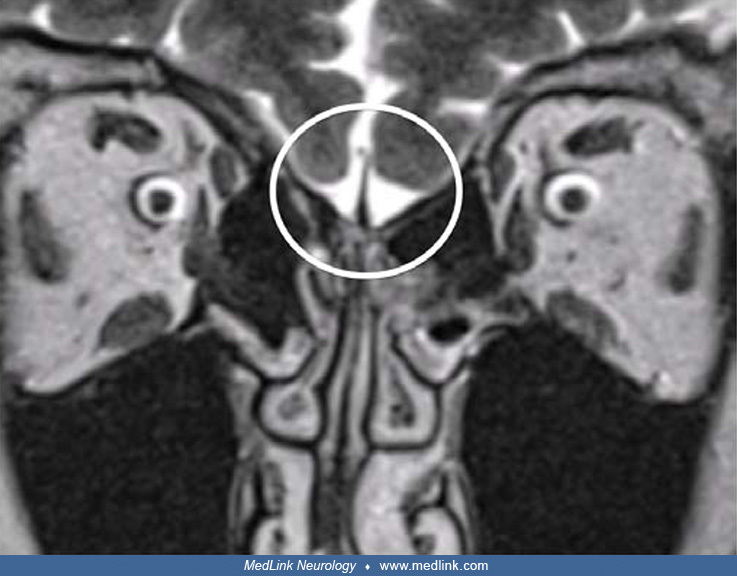
Congenital anosmia may occur with hypoplastic or aplastic olfactory bulbs accompanied by a shallow olfactory sulcus, which is best appreciated on coronal MRI in comparison with normal olfactory bulbs (02; 45). The prevalence of isolated congenital anosmia in the general population is estimated to be 1 in 5000 to 10,000 (45).
Olfactory bulbs are the paired grey rounded structures within the white circled area. (Source: Croy I, Negoias S, Novakova L, Landis BN, Hummel T. Learning about the functions of the olfactory system from people without a sense...
The olfactory bulbs are missing (white circled area). (Source: Croy I, Negoias S, Novakova L, Landis BN, Hummel T. Learning about the functions of the olfactory system from people without a sense of smell. PLoS One 2012;7[3]:e3...
Congenital anosmia is found more often as an isolated symptom but can occur in association with a syndrome, such as Kallmann syndrome. Kallmann syndrome or hypogonadotropic hypogonadism-1 (HH1) with anosmia is most often an X-linked recessive disorder caused by mutation (often a deletion) in the KAL1 gene (ANOS1; OMIM 300836) on chromosome Xp22.3 (although rare autosomal dominant and autosomal recessive forms due to mutations in other genes have been reported). The disorder, with an estimated prevalence of 1 in 84,000 men (77), is characterized by incomplete or absent sexual maturation by the age of 18 years in conjunction with low levels of circulating gonadotropins and testosterone but without other abnormalities of the hypothalamic-pituitary axis and in conjunction with congenital anosmia. In the presence of anosmia, congenital hypogonadotropic hypogonadism is called "Kallmann syndrome"; in the presence of normal olfaction, it is called “normosmic idiopathic hypogonadotropic hypogonadism” (183). About half of the cases of congenital hypogonadotropic hypogonadism are associated with anosmia. Males with Kallmann syndrome show anosmia due to agenesis of the olfactory lobes and hypogonadism secondary to a deficiency of hypothalamic gonadotropin-releasing hormone (149). In a Kallmann fetus, luteinizing hormone-releasing hormone (LHRH)-expressing cells were absent in the brain despite dense clusters of LHRH cells and fibers in the nose (208). LHRH-containing cells and neurites ended in a tangle beneath the forebrain, within the dural layers of the meninges on the dorsal surface of the cribriform plate of the ethmoid bone (208).
Dysosmia in COVID-19. Disorders of the smell and taste are more common among individuals with COVID-19 than among individuals with influenza (30). Chemosensory clinical symptoms are present in at least half of patients with COVID-19 (05; 22; 75; 159; 161; 189).
The risk of COVID-19-associated smell or taste disturbance has progressively fallen with successive waves of infection with the Alpha, Delta, Omicron K, Omicron L, Omicron C, and Omicron B variants, based on their peak intervals (192); consequently, since the Omicron waves, smell and taste disturbances have been of less predictive value in the diagnosis of COVID-19 infection (192; 20).
Chemosensory deficit may be the first symptom ("sentinel symptom") in patients with COVID-19, but there is a wide variation in the proportion of cases in which this is reported to occur (40; 80; 114; 174; 200; 38; 177). A meta-analysis of eight studies, collectively involving 11,054 COVID-19 patients, reported that olfactory and gustatory symptoms appear prior to general COVID-19 symptoms in 65% and 54% of the patients, respectively, based on European, U.S., and Iranian data (203). However, a systematic review of 17 studies found that the onset of dysosmia and dysgeusia occurred 4 to 5 days after other symptoms of the infection and that these chemosensory symptoms typically improved after 1 week, with more significant improvements in the first 2 weeks (204).
Most patients with olfactory and gustatory dysfunction do not present associated nasal congestion or rhinorrhea, and a small group of patients present these alterations in isolation (10; 82; 217). As the disease progresses, patients may experience mild nasal congestion and posterior rhinorrhea, or they may progress without anterior rhinorrhea (87). New-onset chemosensory dysfunction is also common in patients with symptomatic COVID-19 after complete vaccination, and in one study of 153 such patients, approximately half reported associated rhinorrhea, which is a much higher proportion than in unvaccinated cases (226).
Available reports present conflicting data on whether COVID-19 severity is associated with either the degree of olfactory dysfunction or the prognosis for recovery of olfaction, but multiple reports suggest that chemosensory symptoms are most common among those with milder presentations of COVID-19 (28; 46; 181; 205).
Psychophysical testing showed significantly reduced intensity perception and identification ability for both taste and smell functions in patients with COVID-19 (32), although some studies using quantitative testing found olfactory disturbances in nearly all subjects during the acute infection phase, whereas taste or chemesthetic deficits were low (78; 161). Smell and taste loss in COVID-19 are closely associated, although a minority of individuals can experience one or the other (Catton and Gardner 2022a; Catton and Gardner 2022b; 75).
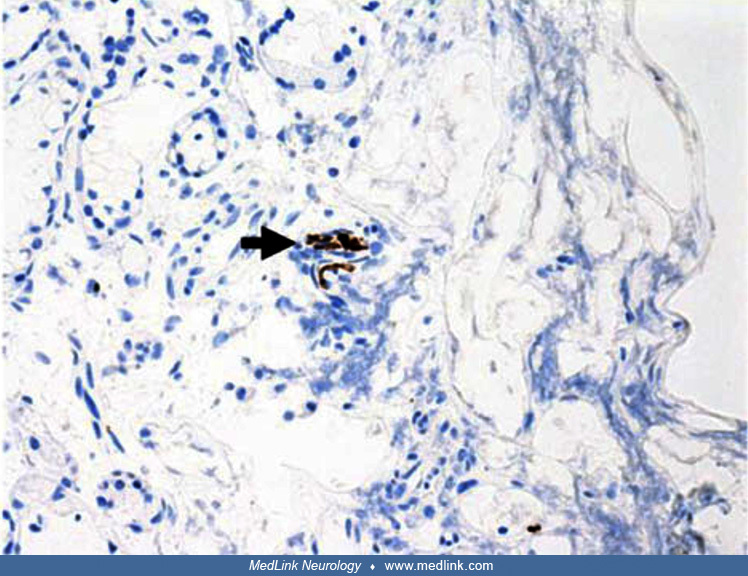
Despite the large number of cases, the pathogenesis of the olfactory dysfunction in COVID-19 has not yet been fully elucidated. A report describing clinical, radiological, and pathological features of a woman who presented with anosmia persisting for more than 3 months after SARS-CoV-2 infection demonstrated significant disruption of the olfactory epithelium from a biopsy, shifting the focus away from invasion of the olfactory bulb and toward the olfactory receptors and the olfactory epithelium (227). Psychophysical tests revealed severe hyposmia and moderate hypogeusia. MRI showed that the olfactory bulb and clefts were of normal volume and were without signal anomalies. She subsequently underwent a biopsy of her left olfactory epithelium. There was extensive loss of surface epithelium, with no associated surface fibrin or inflammatory exudate. Immunohistochemical staining for pancytokeratin AE1/AE3 antibodies demonstrated only focal residual attenuated surface epithelium. There was strong nuclear and cytoplasmic positivity for S100 immunostain in scattered cells within Bowman glands and some small nerve bundles, which is thought to possibly be of trigeminal origin. Immunostaining for the angiotensin-converting enzyme 2 receptor showed focal membrane staining in the S100-positive cells in Bowman glands. There was focal staining for synaptophysin, and neurofilament immunostain highlighted small neurites and nerve bundles in the lamina propria.
MRI in a 63-year-old woman did not reveal any pathological findings; the olfactory bulb and clefts were of normal volume, without signal anomalies. T2-weighted fluid-attended inversion recovery with fat suppression sequence. (S...
MRI in a 63-year-old woman did not reveal any pathological findings; the olfactory bulb and clefts were of normal volume, without signal anomalies. Coronal scans T2-weighted fast spin echo sequence. (Source: Vaira LA, Hopkins C...
Special stain in a 63-year-old woman does not highlight surface basement membrane or inflammatory exudate. (×100) (Source: Vaira LA, Hopkins C, Sandison A, et al. Olfactory epithelium histopathological findings in long-term cor...
Immunostain in a 63-year-old woman showed possible attenuated residual surface epithelial cells, stained brown (arrowhead). (×25) (Source: Vaira LA, Hopkins C, Sandison A, et al. Olfactory epithelium histopathological findings ...
Immunostain in a 63-year-old woman shows strong nuclear and cytoplasmic positivity in scattered cells in structures compatible with Bowman glands (arrow). The same immunostain highlighted small nerve bundles, possibly of trigem...
Immunostaining for angiotensin-converting enzyme 2 (ACE2) receptor in a 63-year-old woman showed focal membrane staining in cells that were also positive for S100 in Bowman glands (arrow). (×200) (Source: Vaira LA, Hopkins C, S...
Focal positive staining for neurofilament immunostain in a 63-year-old woman highlighted small neurites and nerve bundles in lamina propria (arrow). (×100) (Source: Vaira LA, Hopkins C, Sandison A, et al. Olfactory epithelium h...
Chemosensory impairment may be persistent (19; 22; 159; 161; 189), even 1, 2, or 3 years after mild COVID-19 (19; 22; 161; 212), although chemosensory recovery from the Omicron BA.1 subvariant was more favorable than that after the first wave of the pandemic (21). In a prospective observational study, measuring the prevalence of altered sense of smell or taste at follow-up and their variation from baseline, on 403 consecutively assessed adult patients who tested positive for SARS-CoV-2 RNA by polymerase chain reaction during March 2020, 66% reported an altered sense of smell or taste at baseline, whereas 14%, 7%, and 5% reported such alterations at 6 to 24 months, 2 years, and 3 years, respectively (22). Late improvement was possible: of the patients who still experienced smell or taste dysfunction 2 years after COVID-19, 28% and 38% recovered completely and partially, respectively, at the 3-year follow-up (22). Olfactory dysfunction is most likely to persist after 1 year, whereas objectively measured taste dysfunction has typically recovered by that time (212).
Presbyosmia. Presbyosmia (literally “elderly olfaction” or “old age olfaction”) is the gradual loss of olfactory abilities that occurs in most people as they grow older. Clinically significant olfactory loss is common in the elderly but frequently unrecognized, partly because deficits typically accumulate gradually over decades. Indeed, self-reported olfactory impairment significantly underestimates prevalence rates obtained by olfactory testing (165). Because chemosensory impairment is so prevalent among the elderly, many elderly people complain that food lacks flavor, and the elderly account for a disproportionate number of accidental gas poisoning cases (58).
Dysosmia in alpha-synucleinopathies. Olfactory deficits involving odor detection, identification, and discrimination are present in more than 90% of patients with early-stage Parkinson disease (104; 55; 15; 62). A systematic review found that prevalence estimates for olfactory hallucinations in Parkinson disease ranged from 1.6% to 21.0% (225). Occasionally, patients with Parkinson disease may develop pleasant olfactory hallucinations (phantosmias) (126). Impaired olfaction can predate the motor symptoms of Parkinson disease by at least 4 years (198). Idiopathic olfactory dysfunction in first-degree relatives of patients with Parkinson disease is also associated with an increased risk of developing Parkinson disease within 2 to 5 years (178). Olfactory defects in Parkinson disease do not progress markedly with development of motor manifestations (55) and do not correlate well with most other manifestations of the disease (229), except with autonomic defects (84) and cognitive dysfunction, including memory impairment (08). Anosmia in Parkinson disease is associated with autonomic failure, including baroreflex failure and noradrenergic denervation of the heart and other organs, independent of parkinsonism or striatal dopaminergic denervation (84).
Dementia with Lewy bodies is closely allied with both Parkinson disease and Alzheimer disease and is anatomically characterized by the presence of Lewy bodies in both the neocortex and subcortical structures. In dementia with Lewy bodies there is a loss of dopamine-producing neurons in the substantia nigra, similar to that seen in Parkinson disease, and a loss of acetylcholine-producing neurons in the basal nucleus of Meynert, similar to that seen in Alzheimer disease. In dementia with Lewy bodies, as in Parkinson disease, olfactory dysfunction is nearly universal, develops early (before any movement or cognitive disorder), and is often severe (90). Nevertheless, the addition of anosmia to the consensus criteria for dementia with Lewy bodies did not significantly improve overall diagnostic performance (155; 173; 236).
Odor identification is also impaired in patients with REM sleep behavior disorder, a common and very early feature of Lewy body alpha-synucleinopathies (221; 74; 160).
Dysosmia with other neurodegenerative diseases. Although hyposmia is a frequent and early abnormality with alpha-synucleinopathies (ie, Parkinson disease, dementia with Lewy bodies, REM sleep behavior disorder), this is not so with other forms of parkinsonism, including multiple system atrophy, vascular parkinsonism, progressive supranuclear palsy, or corticobasal degeneration, nor is hyposmia a feature of essential tremor (232; 116; 210; 175). Most studies of olfaction in corticobasal degeneration have reported relatively mild deficits, but dysosmia can be moderate or severe in this disorder (175). A mild olfactory loss develops later in multiple system atrophy (116) and is associated with characteristic glial cytoplasmic inclusions in the olfactory bulb and some degree of neuronal loss in the anterior olfactory nucleus; it is unclear if this is of clinical significance (122). Olfactory deficits may also occur with motor neuron disease, but smell testing is not likely to be of clinical value in this condition (67; 90).
Some degree of olfactory loss has also been reported in various other dementing disorders, including Alzheimer disease and frontotemporal dementia (57; 216; 152; 234; 238; 236; 140). Olfactory impairment is more marked early in the course in patients with dementia with Lewy bodies than in those with either Alzheimer disease or frontotemporal dementia (236). Nevertheless, olfactory deficits in Alzheimer disease may be detectable before the appearance of overt memory loss (140), increase with severity of dementia (164; 209; 237), and correlate with the density of neurofibrillary tangles in the entorrhinal cortex and hippocampus (238) and with cortical Lewy body pathology (155). However, it is still unclear whether Alzheimer disease is associated with clinically meaningful hyposmia in the absence of Lewy body pathology (155). Olfactory dysfunction, if apparent in Alzheimer disease, can sometimes help in the differential diagnosis with depression (216; 152). Frontotemporal dementia is also associated with relatively mild olfactory deficits, which are comparable to those seen in Alzheimer disease (154).
For patients with olfactory dysfunction, the prognosis primarily depends on etiology and the degree of residual function but also secondarily on gender, parosmia, smoking habits, and age (106). Male gender, initial presence of parosmia, smoking, and older age are negative prognostic factors (106).
Disorders of the chemosensory senses, smell, and taste are usually less disabling than disorders of the other special senses (vision and hearing). Nevertheless, olfactory impairment significantly contributes to perceived disability and lower quality of life among elderly patients and is a significant predictor of subsequent cognitive decline (158; 165; 238; 207).
Dysosmia in COVID-19. COVID-19-associated chemosensory loss has a substantial negative impact on health-related quality of life beyond mere inconvenience (42; 69). Indeed, altered taste and smell with Covid-19 may cause severe disruption of daily living and quality of life that impacts psychological well-being, physical health, and interpersonal relationships (29; 42; 69). Affected individuals variously reported reduced desire and ability to eat and prepare food; weight gain, weight loss, and nutritional insufficiency; reduced emotional well-being; and impaired intimacy and social bonding (29; 82). "Reduced enjoyment of food" was the most common complaint (87%) in one study (42).
Among healthcare workers who contract COVID-19, olfactory and gustatory loss were associated with emotional distress, anxiety, and depression (63). Moreover, the psychological impact tends to persist even after recovery from the disease, adding to the risk of work-related distress.
The loss of smell and taste improves at a high rate after disease onset in most series (18; 80; 110; 148; 193; 28; 170; 201; 217; 224; 33; 38), but a substantial proportion (approximately 4% to 5%) of patients with Covid-19 might develop long-lasting change in their sense of smell or taste (223). Rapid recovery of olfaction is observed in two thirds of COVID-19-infected people, but the remainder experience a slower pattern of recovery (121). By 1 month, almost all patients have a significant improvement in olfactory threshold and odor discrimination, but not odor identification (110; 170). The recovery of gustation typically occurs before the recovery of olfaction (87). The mean duration of anosmia is 7 days (120), but many of these progress from anosmia to hyposmia and parosmia.
Nevertheless, some series report that between one third and one half of patients have persistent qualitative changes in olfaction (parosmia or phantosmia) that are typically accompanied by qualitative disorders of gustation (parageusia and phantageusia) (163; 73; 81; 100; 211; 144). Some experience long-term deficits with no self-reported improvement at 6 months (73; 81; 99; 100). Persistent parosmia is common, even in those who report at least some recovery of olfactory function (99; 100).
Persistent loss of smell or taste was not associated with persistent SARS-CoV-2 infection (18).
Age under 40 years and the presence of nasal congestion at the time of COVID-19 infection were associated with improved rates of smell recovery, whereas difficulty breathing at the time of COVID-19 infection and prior head trauma were associated with worsened rates of recovery (41).
|
• Olfaction, like disorders of the other special senses, can be conveniently divided into conductive, sensorineural, and central disorders in which (1) conductive disorders involve transmission of the sensory stimuli to the sensory receptors (usually but not always by impeding transmission); (2) sensorineural disorders involve dysfunction of the sensory receptors or conduction of signals from the sensory receptors to the brain; and (3) central disorders involve dysfunction of processing sensory information within the CNS, particularly within the brainstem and cerebrum. | |
|
• Age-related olfactory loss (presbyosmia) is common in the elderly and results from normal aging, certain diseases (especially Parkinson disease and dementia with Lewy bodies), medications, surgical interventions, and prior environmental exposures. | |
|
• The elderly have higher olfactory thresholds, perceive suprathreshold odors less intensely, and are less able to discriminate odors or to recognize and identify common odors. | |
|
• COVID-19-related chemosensory dysfunction primarily results from a loss of function of olfactory sensory neurons and tastebuds that is mainly caused by infection, inflammation, and subsequent dysfunction of supporting non-neuronal cells in the mucosa. | |
|
• A multi-ancestry study of COVID-19-related chemosensory dysfunction from 69,841 individuals identified a genome-wide significant locus in the vicinity of the UGT2A1 and UGT2A2 genes; both genes are expressed in the olfactory epithelium and play a role in metabolizing odorants. | |
|
• Sudden dysosmia in COVID-19 is not related to central involvement due to neuroinvasive SARS-CoV-2. Instead, it is associated with subtle cerebral metabolic changes in core olfactory and high-order cortical areas resulting from deafferentation and active functional reorganization secondary to the lack of olfactory stimulation. | |
|
• Olfactory loss in Parkinson disease and dementia with Lewy bodies is not due to damage to the olfactory epithelium but, instead, results from CNS abnormalities. |
The nasopharynx and the olfactory pathways. Olfaction is the sensation of smell that results from detecting odorous substances aerosolized in the environment.
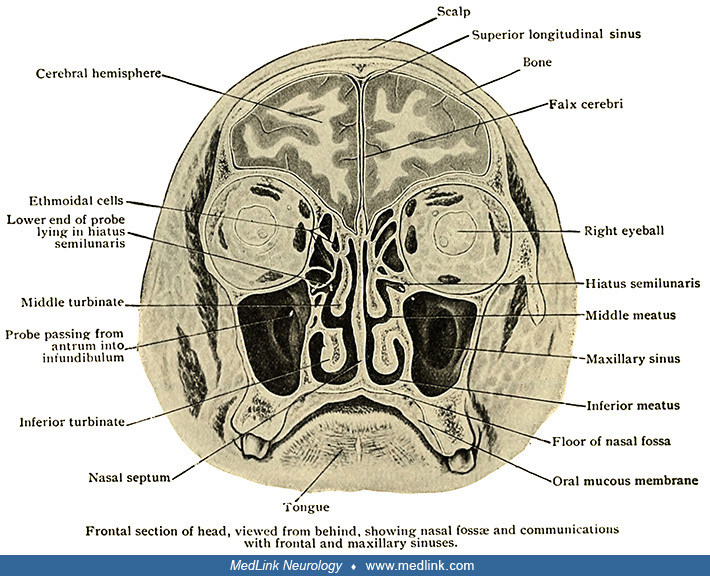
The nasal passages are divided in the midline by the nasal septum. Each lateral nasal wall is formed by several turbinates, spongy curled bones covered by mucosa. The turbinates protrude into the nasal passages and serve to humidify, warm, and cleanse air passing through the nasal passages to the lungs. The irregular path of airflow through the nose results in turbulence, which helps direct air and odorant molecules superiorly toward the olfactory epithelium, facilitating olfaction. The nasal passages also provide openings into various air spaces: the frontal, maxillary, ethmoid, and sphenoid sinuses; the sphenoethmoidal recess; and the middle ear (via the Eustachian tube).
The olfactory receptor cells are in a patch of specialized epithelium--the olfactory epithelium--that straddles the crest of the nasal vault on each side. In two young men, German anatomist Albert von Brunn (1849-1895) (his full name was Ferdinand Albert Wilhelm von Brunn) determined the extent of the olfactory epithelium to be approximately 50 cm2, which led to the eponym "Brunn membrane" for the olfactory epithelium (26). Brunn elaborated the histology of the olfactory epithelium and teased apart the component cells, distinguishing particular olfactory receptor cells and supporting cells. Olfactory receptor cells are bipolar neurons characterized by a tapered shape and the presence of cilia protruding into the nasal vault, where they can detect odorant stimuli that have dissolved in the nasal mucus. Olfactory receptor neurons are activated when airborne molecules in inspired air bind to olfactory receptors expressed on their cilia. Olfactory cilia are constantly replaced, an ability not characteristic of the other sensory receptors.
(Source: Romano N, Federici M, Castaldi A. Imaging of cranial nerves: a pictorial overview: insights Imaging. 2019;10[1]:33. Creative Commons Attribution 4.0 International [CC BY] license, creativecommons.org/licenses/by/4.0.)<...
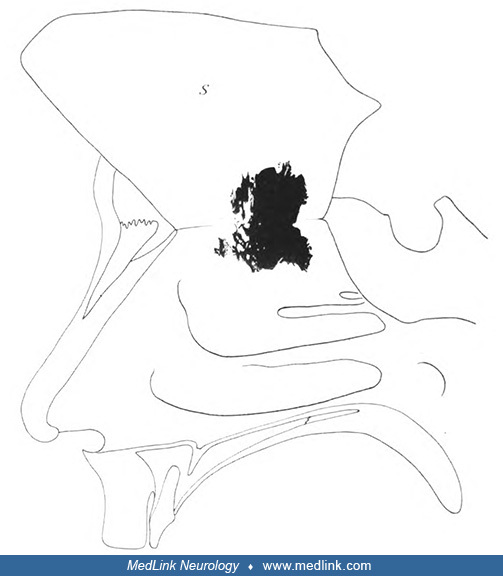
"Right nasal cavity. The septum S detached all around, with the exception of the upper edge, and folded upwards. The dark figure represents the spread of the olfactory epithelium as it appears after reconstruction ... male, 40 ...
"Right nasal cavity. The septum S detached all around, with the exception of the upper edge, and folded upwards. The dark figure represents the spread of the olfactory epithelium as it appears after reconstruction ... male, 30 ...
The olfactory epithelium is a pseudostratified epithelium composed mainly by sustentacular cells, globose basal cells, horizontal globose cells, and both immature and mature olfactory sensory neurons; the olfactory ensheathing cells are an important cell type populating the submucosa (124).
In the nose, mature olfactory sensory neurons expressing the same odorant receptors gene are stochastically distributed within a spatially restricted area of the olfactory epithelium, also known as a “zone.” Early studies identified four non-overlapping odorant receptor expression zones, but later studies identified 9 to 12 partially overlapping zones (124).
Each mature olfactory sensory neuron expresses one allele of a single odorant receptor gene: the "one neuron - one receptor" rule (124).
Most intact odorant receptors are expressed in the olfactory epithelium across a large dynamic range, with only a minority being expressed at very high levels (124).
Odorants are detected by odorant receptors in a combinatorial fashion: one odorant can activate multiple odorant receptors, and each odorant receptor can detect more than one odorant. Recent studies that analyzed the responses of odorant receptors to specific odorants presented as part of odor mixtures found that odorants, in addition to their agonist role, can also modulate odorant receptor activity, serving as antagonists, inverse agonists, partial agonists, and synergistic ligands (124).
An inverse agonist binds to the same receptor as an agonist but brings about the opposite response to that of an agonist, whereas an antagonist binding to such a receptor will disrupt the interaction and the function of both the agonist and the inverse agonist at the receptor.
The olfactory receptors (also known as odorant receptors) on the cilia of the olfactory receptor neurons belong to a G-protein-coupled receptor superfamily (specifically the class A rhodopsin-like family of G protein-coupled receptors). The olfactory receptors form a multigene family consisting of around 800 genes in humans.
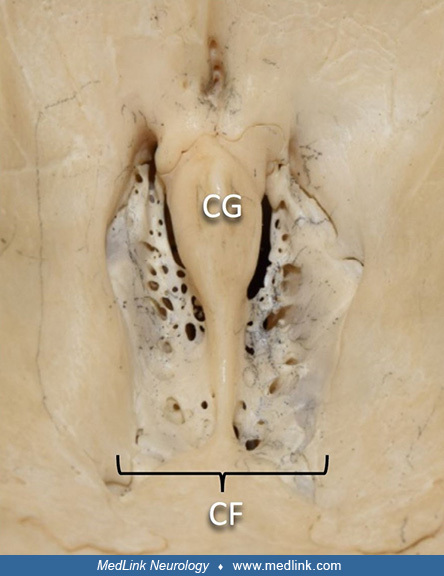
The neural processes of the bipolar olfactory receptor neurons pass upward through small holes in the cribriform plate of the ethmoid bone on the cranial floor of the anterior fossa.
Legend: (CG) crista galli, (CF) cribriform plate. (Source: Edwards B, Wang JM, Iwanaga J, Loukas M, Tubbs RS. Cranial nerve foramina part I: a review of the anatomy and pathology of cranial nerve foramina of the anterior and mi...
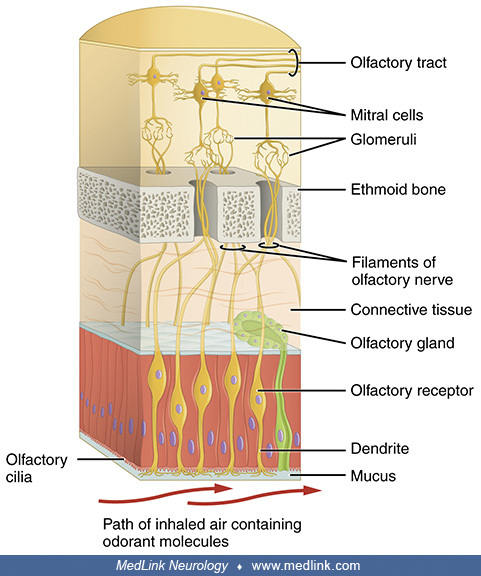
The (olfactory) glomeruli are spherical structures in the olfactory bulb where synapses form between the terminals of the olfactory nerve and the dendrites of mitral, periglomerular, and tufted cells. All glomeruli are located near the surface of the olfactory bulb. A glomerulus is made up of a globular tangle of axons from the olfactory receptor neurons and dendrites from the mitral and tufted cells of the olfactory bulb. Each glomerulus is surrounded by various juxtaglomerular neurons (eg, periglomerular, short axon, and external tufted cells) and astrocytes. In humans, there are roughly 1100 to 1200 glomeruli, but the number decreases with age; few remain by the age of 80.
(Source: OpenStax May 18, 2016. Version 8.25 from the Textbook OpenStax Anatomy and Physiology. Creative Commons Attribution 4.0 International [CC BY 4.0] license, creativecommons.org/licenses/by/4.0. Labeling modified...
Each olfactory receptor neuron expresses one olfactory receptor, which responds to different odorants. Odorant molecules bind to olfactory receptors on cilia. Olfactory receptors activate olfactory receptor neurons, which trans...
The olfactory bulb and the olfactory tract lie in the olfactory sulcus on the underside of the frontal lobe, between the gyrus rectus and the orbital frontal gyri. The olfactory tract divides at the olfactory trigone into medial, middle, and lateral roots. The entire length from the olfactory bulb to the trigone rests above the skull floor of the anterior fossa, at the level of the bridge of the nose when projected from outside the skull. Although located just lateral to the midline, they, nevertheless, are anterior to the midline chiasm and infundibulum.
The mechanisms responsible for the detection of odors and the formation of a "smell map" were delineated by groundbreaking work done by Richard Axel at Columbia University and Linda Buck at the Fred Hutchinson Cancer Research Center in Seattle, who shared the 2004 Nobel Prize in Physiology or Medicine “for their discoveries of odorant receptors and the organization of the olfactory system.”
Types of olfactory disorders. Olfaction, like disorders of the other special senses, can conveniently be divided into conductive, sensorineural, and central disorders, whereby (1) conductive disorders involve transmission of the sensory stimuli to the sensory receptors (usually but not always by impeding transmission); (2) sensorineural disorders involve dysfunction of the sensory receptors or conduction of signals from the sensory receptors to the brain; and (3) central disorders involve dysfunction of processing sensory information within the CNS, particularly within the brainstem and cerebrum (128).
Conductive olfactory disturbances. Odorants reach the sensory receptors in the olfactory neuroepithelium by two pathways: orthonasally through the nostrils or retronasally through the nasopharynx (64). With retronasal olfaction, odorants are delivered in a liquid or semiliquid phase during eating or drinking, are volatilized, and are then combined with gustatory and somatosensory sensations to form a composite sensation of flavor. This process requires adequate mastication to release volatile odorants and sufficient mouth and swallowing movements to effectively pump the odorants retronasally.
Conductive (or “transport”) olfactory problems impede the passage of odorants either orthonasally or retronasally and generally cause hyposmia (rather than anosmia) because the obstruction is usually incomplete. Some common problems that produce conductive olfactory disturbances include upper respiratory infections, chronic rhinosinusitis, and nasal polyposis. In addition, elderly patients with complete or palate-covering dentures have lower olfactory sensitivity than those who are dentate or who wear dentures that do not cover the palate. This is due, in part, to impairment with chewing and mouth movements and interference with the movement of odorants retronasally (a conductive defect) (64).
The mechanisms of olfactory dysfunction in rhinosinusitis are not limited to conductive olfactory problems, however. Indeed, chronic inflammation in the olfactory epithelium impairs not only olfactory sensory neurons but also nonneuronal cells that are responsible for regeneration and support for neurons, implying a sensorineural olfactory component (219).
Sensorineural olfactory disturbances. The olfactory receptors are in the olfactory neuroepithelium on the superior-nasal septum and lateral wall of the nasal cavity. The dendritic ends of these bipolar sensory cells project into the overlying nasal mucous, whereas the unmyelinated axons project through the cribriform plate of the ethmoid bone via small bundles that comprise the filaments of the short olfactory nerve to ultimately synapse in the olfactory bulb.
With sensorineural olfactory problems, the odorants contact the olfactory receptors of the bipolar neurons in the olfactory neuroepithelium, but dysfunction of these components prevents the information from being processed. Sensorineural olfactory problems can be caused by head trauma, tobacco smoking, other toxins, drugs, nutritional disorders (including zinc deficiency, vitamin A deficiency, cobalamin deficiency), influenza-like viral infections, various comorbid medical conditions (hypothyroidism, diabetes, Sjogren syndrome, renal failure, and liver disease, including cirrhosis), and structural lesions involving the area of the cribriform plate (such as subfrontal meningioma).
Moderate to severe traumatic brain injury is associated with prominent olfactory dysfunction (222). Mechanisms of posttraumatic olfactory dysfunction include direct injury to the olfactory epithelium (causing a sensory olfactory deficit), shearing effect on the fragile olfactory fibers at the cribriform plate (causing a neural deficit), or potential brain contusion or intraparenchymal hemorrhage (causing a central olfactory deficit). Posttraumatic phantosmia is likely due to release hallucinations after shearing injury to the fragile olfactory nerves passing through the cribriform plate (128; 09).
Hyposmia is common in smokers, occurs in a dose-dependent fashion, and apparently results from increased death of olfactory sensory neurons, which eventually overwhelms the regenerative capacity of the olfactory neuroepithelium (44; 228; 106).
Hyposmia is common in hypothyroidism, occurring in about 20% of cases, and contributes to anorexia and lack of interest in eating, but it largely reverses with replacement hormone therapy (153).
In the absence of local nasal disease, unilateral anosmia suggests a structural lesion of the olfactory nerve filaments, bulb, tract, or stria. An olfactory groove meningioma, for example, may cause ipsilateral sensorineural anosmia in conjunction with optic atrophy in the ipsilateral eye (due to optic nerve compression) and papilledema in the contralateral eye (due to increased intracranial pressure), a constellation of manifestations designated eponymically as Foster-Kennedy syndrome.
Central olfactory disturbances. As recognized very early by Spanish histologist Ramón y Cajal, the central olfactory pathways are complex and involve many relay centers and circuits (76; 39).
The olfactory bulb may constitute the “olfactory thalamus” (although it has also been suggested more provocatively that the olfactory bulb may instead serve as the primary olfactory cortex) (89; 118; 12). Second-order neurons from the olfactory bulb travel posteriorly as the optic tract in the olfactory sulcus on the orbital surface of the frontal lobes. The optic tracts divide into medial and lateral striae, with fibers from the medial striae decussating in the anterior commissure to terminate in the contralateral cerebral hemisphere, whereas fibers from the lateral striae project to the ipsilateral primary olfactory cortex, amygdala, septal nuclei, and hypothalamus. Because of the bilateral cortical representation for smell in the piriform cortex, unilateral lesions distal to the decussation of the olfactory fibers generally do not cause clinically important olfactory dysfunction, although deficits in olfactory discrimination and detection may be detected with major unilateral damage to the frontal or temporal lobes. Disorders that can interfere with central olfaction include epilepsy, head injury (particularly with contusion of the temporal tips against the anterior portion of the middle fossa), and various neurodegenerative conditions (such as Parkinson disease, dementia with Lewy bodies, and Alzheimer disease). Some central disorders, such as epilepsy, produce irritative phantosmias, whereas others that interfere with the olfactory pathways may produce release hallucinations.
An interesting example of olfactory aura (irritative phantosmia) was reported by Rao and colleagues (188). A 47-year-old man presented with the subacute onset of constitutional symptoms, diffuse lymphadenopathy, and stereotyped spells involving an unpleasant olfactory aura followed by nausea, dizziness, disorientation, and unresponsiveness. A routine electroencephalogram was normal, and he was treated empirically with levetiracetam without benefit. Further evaluation disclosed lymphadenopathy on chest CT, focal dyscognitive seizures of temporal lobe origin, CSF with lymphocytic pleocytosis, hyponatremia, and serum positive for voltage-gated potassium channel antibodies, consistent with limbic encephalitis. Lymph node biopsy revealed multicentric Castleman disease. His symptoms improved with antiepileptic drugs and immunotherapy.
EEG in a 47-year-old man during a seizure (red bar) arising from light sleep. Initially, there is emergence of theta/alpha range frequencies broadly over the left temporal region, maximal at electrodes F7 and T3. Clinically, th...
EEG in a 47-year-old man during a seizure (red bar) arising from light sleep. Left temporal waveforms become rhythmic. Clinically, the patient was observed to have lip-smacking automatisms during this seizure. (Source: Rao VR, ...
EEG in 47-year old man during a seizure (red bar) arising from light sleep. Left temporal waveforms evolve in frequency and amplitude while acquiring sharpened elements. Clinically, the patient was observed to have lip-smacking...
EEG in a 47-year-old man shows interictal background after a seizure. (Source: Rao VR, Lim LE, Fong D, Garga NI, Parko KL. Multicentric Castleman's disease with voltage-gated potassium channel antibody-positive limbic encephali...
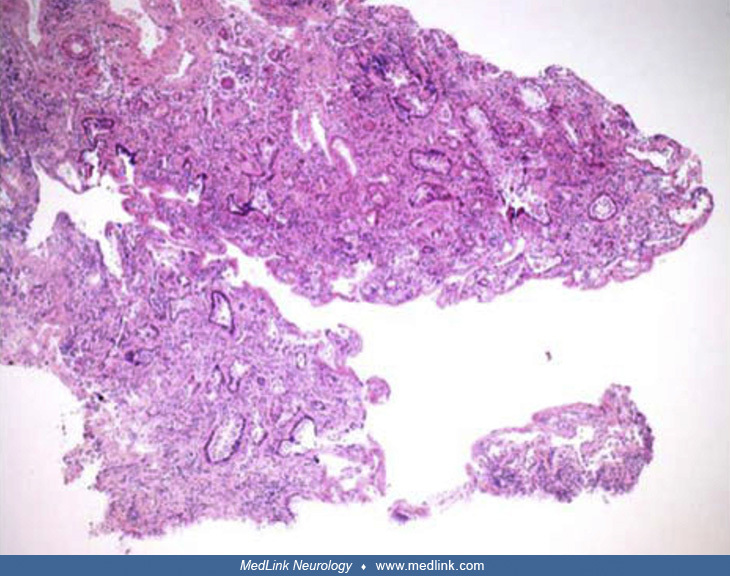
Lymph node biopsy pathology (10x) in a 47-year-old man with subacute onset of constitutional symptoms, diffuse lymphadenopathy, and stereotyped spells involving olfactory aura, nausea, disorientation, and unresponsiveness. Atro...
Lymph node biopsy pathology (20x) in a 47-year-old man with subacute onset of constitutional symptoms, diffuse lymphadenopathy, and stereotyped spells involving olfactory aura, nausea, disorientation, and unresponsiveness. Germ...
Presbyosmia. Age-related olfactory loss (presbyosmia) is common in the elderly and results from normal aging, certain diseases (especially Parkinson disease and dementia with Lewy bodies), medications, surgical interventions, and prior environmental exposures (58; 53; 206; 72; 165; 145; 190). The components related to aging, per se, are relatively small (but significant), whereas most of the age-related functional declines of the chemosenses (ie, olfaction and gustation) are attributable to accumulated insults to the sensory system, smoking, medications, and comorbid disease (165; 145).
The elderly have higher olfactory thresholds, perceive suprathreshold odors less intensely, and are less able to discriminate odors or to recognize and identify common odors (31; 206; 56). Part of the decline in olfactory abilities with age results from degeneration of the olfactory bulb. The number of mitral cells and glomeruli in the olfactory bulb declines markedly with age, at an approximate rate of 10% per decade, so that less than 30% of these elements remain by the ninth and tenth decades (156). Functional imaging shows significantly lower activation in brain regions receiving primary olfactory projections (piriform cortex, entorhinal cortex, and amygdala) among the elderly (35).
A systematic review of associations of olfactory function with brain structural and functional outcomes in older adults without and with cognitive impairment found that lower olfactory function was associated with smaller volumes in the temporal lobe (hippocampus, parahippocampal gyrus, fusiform gyrus), olfactory-related regions (piriform cortex, amygdala, entorhinal cortex), pre- and postcentral gyri, and globus pallidus (24).
Dysosmia in COVID-19. Severe acute respiratory syndrome coronavirus-2 (SARS-CoV-2) enters human host cells via mechanisms facilitated by angiotensin-converting enzyme 2 (ACE2) and transmembrane protease serine 2 (TMPRSS2). SARS-CoV-2 infection can damage the olfactory receptor cells in the nasal neuroepithelium or the olfactory bulb (119; 37; 101).
(Source: Hu B, Gong M, Xiang Y, Qu S, Zhu H, Ye D. Mechanism and treatment of olfactory dysfunction caused by coronavirus disease 2019. J Transl Med 2023;21[1]:829. Creative Commons Attribution 4.0 International [CC BY 4.0] lic...
(Source: Hu B, Gong M, Xiang Y, Qu S, Zhu H, Ye D. Mechanism and treatment of olfactory dysfunction caused by coronavirus disease 2019. J Transl Med 2023;21[1]:829. Creative Commons Attribution 4.0 International [CC BY 4.0] lic...
(a) Supporting cells affect olfactory sensory neurons functions through tight junctions. (b) SARS-CoV-2 enters and damages olfactory sensory neurons. (c) Amplification of inflammatory factors leads to olfactory sensory neuron i...
COVID-19-related chemosensory dysfunction primarily results from a loss of function of olfactory sensory neurons and taste buds, mainly caused by infection, inflammation, and subsequent dysfunction of supporting non-neuronal cells in the mucosa (150). A multi-ancestry study of COVID-19-related chemosensory dysfunction from 69,841 individuals identified a genome-wide significant locus in the vicinity of the UGT2A1 and UGT2A2 genes; both genes are expressed in the olfactory epithelium and play a role in metabolizing odorants (213; 101). Proposed mechanisms for olfactory dysfunction in COVID-19 include: (1) the infection of sustentacular cells, which triggers olfactory dysfunction; and (2) the inflammatory reaction, which includes the amplification of inflammatory factors, damages the nasal epithelium, and leads to olfactory sensory neuron impairment, likely prolonging and exacerbating early olfactory dysfunction (37; 101). Sustentacular cells are likely to be the entry point of SARS-CoV-2 (rather than olfactory neurons) and are more susceptible to early injury (37).
A multi-ancestry study of COVID-19-related chemosensory dysfunction from 69,841 individuals identified a genome-wide significant locus in the vicinity of the UGT2A1 and UGT2A2 genes; both genes are expressed in the olfactory epithelium and play a role in metabolizing odorants (213).
Early studies using MRI demonstrated that dysosmia in COVID-19 may be associated with a reversible obstruction of the olfactory clefts due to transient edema (68).
Structural brain MR and positron emission tomography with [18F]-fluorodeoxyglucose (FDG-PET) were acquired simultaneously on a hybrid PET-MR in 12 confirmed cases of COVID-19 with complaints of sudden dysosmia (168). Sudden dysosmia in COVID-19 was not related to central involvement due to neuroinvasive SARS-CoV-2 but was instead associated with subtle cerebral metabolic changes in core olfactory and high-order cortical areas resulting from deafferentation and active functional reorganization secondary to the lack of olfactory stimulation (168).
Interleukin 6 (IL-6), a proinflammatory cytokine that is promptly but transiently produced in response to infections and tissue injuries, contributes to host defense through the stimulation of acute phase responses, hematopoiesis, and immune reactions. The rapid recovery of smell and taste functions in patients with COVID-19 may, in part, be attributable to changes in IL-6 levels rather than to recovery from viral injury to neurons (34; 217).
A high proportion of patients with COVID‐19 describe a loss of taste, but standardized functional testing of the gustatory modalities of sweet, sour, salty, and bitter often does not reveal a bona fide hypogeusia (97). Therefore, subjectively altered taste is probably caused, in part, by impaired retronasal olfaction (97; 235).
Dysosmia in alpha-synucleinopathies. Olfactory loss in Parkinson disease and dementia with Lewy bodies is not due to damage to the olfactory epithelium but, instead, results from CNS abnormalities (103; 239; 08). Pathology of the olfactory bulb and tract occurs before motor signs of Parkinson disease (103). In alpha-synucleinopathies, olfactory dysfunction relates specifically to Lewy body pathology and correlates with cardiac sympathetic denervation, independently of striatal dopamine deficiency or parkinsonism (83). Impaired olfaction in Parkinson disease is associated with the presence of Lewy bodies and neuronal loss in the olfactory bulb and tract, with a strong correlation between neuronal loss and disease duration (176). In Parkinson disease, the olfactory bulb contains numerous Lewy bodies, and severe neuronal loss is present in the anterior olfactory nucleus (122). Immunolabeling for alpha-synuclein in the olfactory bulb and tract occurs prior to clinical signs of parkinsonism in dementia with Lewy bodies (103). The presence of Lewy body alpha-synucleinopathy in the olfactory bulb accurately predicts the presence of Lewy body pathology in other brain regions (11).
The pathophysiology of hyposmia in Lewy body alpha-synucleinopathies is not well understood and may have multiple components, including those resulting from degenerative changes in the olfactory bulb and primary olfactory cortex, as well as limbic dysfunction and possibly prefrontal dysfunction (104; 16; 233; 17; 08). Functional imaging indicates that reduced neuronal activity in the amygdala, hippocampus, and piriform cortex (uncus) contributes to olfactory dysfunction in Parkinson disease (233; 08). Hyposmia in Parkinson disease is more closely associated with cholinergic denervation of limbic archicortex (hippocampus and amygdala) than with nigrostriatal dopaminergic denervation (17). However, because olfactory threshold and odor identification in Parkinson disease are not related to duration of disease, to current therapy with levodopa or anticholinergic drugs, or to “on” and “off” states, olfactory impairment in Parkinson disease likely involves mechanisms that are not due to dopaminergic or cholinergic denervation and that are not influenced by pharmacologic manipulation of dopaminergic or cholinergic status (182).
|
• Olfactory senescence starts by the fifth decade and accelerates with advancing years, preferentially involving pleasant odors. | |
|
• Smell identification ability markedly declines after the seventh decade; major olfactory impairment is present in about one third to one half of those aged 65 to 80 and in two thirds to four fifths of those over the age of 80. | |
|
• There is a high prevalence of olfactory and gustatory dysfunction among patients infected with novel COVID-19. | |
|
• Chemosensory disorders are relatively uncommon in pediatric cases of COVID-19 compared with adults. |
Olfactory senescence starts by the fifth decade and accelerates with advancing years, preferentially involving pleasant odors (58; 90; 56). Almost two thirds of the patients aged 80 and older have olfactory impairment (165). Male gender, current smoking, medications, cumulative exposure to toxic fumes, prior head trauma, and comorbid conditions (such as nasal congestion, upper respiratory tract infection, sinusitis, systemic viral illness, epilepsy, cerebrovascular disease, and neurodegenerative diseases) are associated with an increased prevalence of olfactory impairment in the elderly (206; 72; 165; 167).
Smell identification ability declines markedly after the seventh decade; major olfactory impairment is present in about one third to one half of those aged 65 to 80 and in two thirds to four fifths of those over the age of 80 (58; 165; 56; 125).
Dysosmia in COVID-19. There is a high prevalence of olfactory and gustatory dysfunction among patients infected with novel COVID-19 (03; 110; 07; 13; 201; 23; 88; 113; 142). Dysosmia is more common in patients with COVID-19 than in patients suffering from other viral infections or controls (179). In a meta-analysis of 24 studies collectively involving 8438 patients with test-confirmed COVID-19 infection from 13 countries, the pooled proportions of patients presenting with olfactory dysfunction and gustatory dysfunction were 41% and 38%, respectively (03). In a separate meta-analysis of eight studies collectively involving 11,054 COVID-19 patients, dysosmia and dysgeusia symptoms were present in 75% and 81% of cases of COVID-19, respectively (203). The prevalence of olfactory and gustatory dysfunction is negatively correlated with age in adults (03; 87; 138; 108; 172). The estimated prevalence of olfactory dysfunctions is higher with objective measurements than with self-reports (03; 141).
Chemosensory disorders are relatively uncommon in pediatric cases of COVID-19 compared with adults (71; 123; 180). In one series, only 6% of 122 pediatric COVID-19 cases had a chemosensory disorder, and all of the affected children fully recovered by the end of the third week of illness, even though one showed restricted diffusion in the splenium of the corpus callosum (71). In another series, 28% of 141 adolescents had chemosensory dysfunction, with a self-reported duration of 2 to 15 days (mean 5.7 days) (123).
The severity of COVID-19-associated chemosensory dysfunction has dropped significantly with the advent of the Omicron variant (23). Some studies also reported that the frequency of chemosensory dysfunction also declined with the advent of the Omicron variant (23), whereas others found no change in the prevalence of chemosensory dysfunction during the period of Omicron dominance (218; 230). In one study in the Omicron era, individuals with chemosensory dysfunction had significantly higher rates of influenza-like upper respiratory symptoms, xerostomia, and halitosis (230); moreover, olfactory dysfunction was a risk factor for the prevalence of taste dysfunction in patients with Omicron infection.
For most olfactory disorders, prevention opportunities are limited.
Measures to decrease traumatic brain injury would help significantly to prevent traumatic dysosmias, including measures to decrease sports-related brain injuries (American football, boxing, mixed martial arts, and heading soccer balls), encourage or require motorcycle helmets, limit alcoholism, and prevent falls. Prevention of COVID-19-related chemosensory dysfunction is feasible with vaccination, although vaccinated individuals still carry a risk of developing COVID-19 and related chemosensory dysfunction (226).
Patients and clinicians frequently overlook olfactory disorders, and many patients seem genuinely unaware of their deficits. Therefore, it is particularly useful to screen for olfactory dysfunction in certain patient populations, including those who have or may have suffered a significant head injury and those suspected of having a synucleinopathy.
Many individuals with olfactory disorders come to medical attention because patients note a lack of taste in their food rather than a lack of smell. Olfaction should routinely be assessed in patients who complain of impaired olfaction or impaired gustation.
|
• Medications that can alter olfactory function are numerous and varied and include levodopa, bromocriptine, lithium, opiates, various lipid-lowering drugs, calcium-channel blockers, beta-blockers, antimicrobials, and antineoplastics. | |
|
• The University of Pennsylvania Smell Identification Test (UPSIT), the most often used commercial odor identification test, may be self-administered in an office environment and scored by a medical assistant or nurse. | |
|
• Sudden onset of dysosmia during the COVID-19 pandemic generally does not require extensive investigation. | |
|
• Markedly reduced olfaction in a parkinsonian patient is supportive of Parkinson disease or dementia with Lewy bodies. |
Review medications for medication-induced dysosmia. Medications that can alter olfactory function are numerous and varied and include levodopa, bromocriptine, lithium, opiates, various lipid-lowering drugs, calcium-channel blockers, beta-blockers, antimicrobials, and antineoplastics (206; 220). Some medications interfere with the process of sensory transduction (eg, by interfering directly with the receptor or its components, such as G-proteins, adenylate cyclase, or receptor kinase), others interfere with neurotransmitters involved in olfactory processing, and still others are directly toxic to olfactory neuroepithelium or the nerves themselves. For example, chemotherapy agents cause hyposmia by direct toxicity to mucosa and nerves and by inhibition of mucosal cell growth and replacement. In addition, zinc-induced anosmia due to necrosis of nasal neuroepithelium occurred with intranasal application of zinc gluconate gel in commercial cold preparations, but this formulation has been withdrawn (04; 215).
A straightforward framework for diagnostic evaluation of parosmia has been proposed (39).
Physical examination and office testing. Physical examination should include examination of the nose, mouth, and throat as well as neurologic examination.
Office testing of smell can include well-standardized, commercially available tests (such as the University of Pennsylvania Smell Identification Test, or UPSIT) or crude approaches utilizing identification of a few readily available odorants (such as oil of wintergreen or oil of cloves) (59; 60; 54). More complicated odor identification and detection tests are also available but are rarely practical outside specialized diagnostic laboratories (54).
The University of Pennsylvania Smell Identification Test (UPSIT), the most often used commercial odor-identification test, is a forced-choice olfactory discrimination test that uses microencapsulated odorants in standardized “scratch and sniff” booklets. It has now been translated into several languages and is widely employed due to its accurate categorization of respondents and its ease of use, with no need for complex equipment. Patients are instructed to "scratch and sniff" each test odorant in turn and then select what they believe is the best choice for each one from among four possible odor identifications. The test can be self-administered, and because of the clever design of the booklets, it can be rapidly scored by a medical assistant or nurse. The correct responses are summed across all four booklets, providing a total score ranging from 0 to 40. Age- and gender-specific norms are provided that allow categorization into probable malingering, anosmia, various degrees of hyposmia (ie, mild, moderate, and severe "microsmia"), and normosmia. In actuality, the test will correctly identify on some malingerers.
Irritant substances, such as ammonia, are sometimes employed when psychogenic or malingered anosmia is a consideration because such substances are perceived via trigeminal afferent pathways instead of through the olfactory system.
Imaging. Cranial imaging (CT and MRI) is most useful to detect abnormalities in the nasal cavities (eg, nasal polyposis), nasal sinuses (eg, sinusitis), and anterior cranial fossa (eg, meningioma) in patients with olfactory disorders.
Biopsy of olfactory epithelium. Success rates of obtaining olfactory epithelium in biopsies performed from the nasal septum vary between 40% to 89%, and approximately half have material of sufficient quality for morphological characterization (79).
Olfactory biopsies from the superior part of the nasal septum did not significantly affect smell capacity (79). These biopsies obtain high rates of olfactory neurons (approximately 90%) (79).
Dysosmia in COVID-19. Sudden onset of dysosmia during the COVID-19 pandemic generally does not require extensive investigation. New loss of smell (anosmia/hyposmia) is now recognized as a COVID-19-related symptom (119; 148; 162; 194; 195; 202; 171). Indeed, dysosmia is the best predictor of COVID-19 positivity amongst those with symptoms of respiratory illness (194; 81). There is a significant and strong association between positive real-time polymerase chain reaction (RT-PCR) test results for SARS-CoV-2 and subjective or objectively demonstrated hyposmia (148; 194; 202). The combination of subjective hyposmia and cough significantly increases the odds of having a positive RT-PCR (148), as does the combination of hyposmia and fever (195). The prevalence of olfactory loss is so high among people with COVID-19 that a person who has normal olfaction on formal testing is unlikely to be infected with SARS-CoV-2 (91). During the COVID-19 pandemic, olfactory dysfunction may be an early sign of COVID-19, even without other upper respiratory infection or otolaryngologic symptoms (115). Given the high prior probability of COVID-19 and the strong association of COVID-19 with new-onset chemosensory dysfunction, new-onset chemosensory dysfunction has been demonstrated to be highly predictive for positive RT-PCR test results for SARS-CoV-2.
Dysosmia in parkinsonism. Markedly reduced olfaction in a parkinsonian patient is supportive of Parkinson disease or dementia with Lewy bodies (232), whereas normal smell identification is rare with these conditions and should prompt a review of the diagnosis (unless the patient is female with tremor-dominant disease) (90). Preserved or mildly impaired olfactory function in a parkinsonian patient is more likely to occur with atypical parkinsonism, such as vascular parkinsonism, multiple system atrophy, progressive supranuclear palsy, or corticobasal degeneration (117; 90). However, these heuristic clinical rules are not absolute; some patients with corticobasal degeneration, for example, have moderate or severe impairment (175).
|
• Anosmia associated with chronic allergic rhinitis, nasal polyposis, and chronic sinusitis may respond to glucocorticoids, especially if administered systemically. | |
|
• Olfactory training is recommended for all patients with persistent COVID-19-related dysosmia of more than 2 weeks’ duration. | |
|
• Olfaction in patients with Parkinson disease does not improve with levodopa therapy. |
Anosmia associated with chronic allergic rhinitis, nasal polyposis, and chronic sinusitis may respond to glucocorticoids, especially if administered systemically. Results with zinc sulfate therapy, even in patients with documented zinc deficiency, have been mixed.
Dysosmia in COVID-19. Consensus guidelines of the British Rhinological Society for the management of new-onset loss of sense of smell during the COVID-19 pandemic include the following: (1) olfactory training was recommended for all patients with persistent loss of sense of smell of more than 2 weeks’ duration; (2) oral steroids, steroid rinses, and omega 3 supplements may be considered; (3) alpha-lipoic acid was not recommended; and (4) because of panel disagreement, no recommendation was made regarding using Vitamin A drops (98).
Corticosteroids. The literature concerning topical corticosteroids for COVID-19 olfactory dysfunction is at best inconsistent, even in systematic reviews and meta-analyses. Indeed, one systematic review and metaanalysis found that although nasal steroid therapy does not result in significant adverse effects, it is ineffective in the treatment of COVID-19 olfactory dysfunction (36), whereas another systematic review and meta-analysis concluded that "topical corticosteroid treatment is a viable option for improving COVID-19-related olfactory dysfunction" (231). Mometasone furoate nasal spray as a topical corticosteroid for the treatment of post-COVID-19 anosmia is not superior to olfactory training regarding smell scores, duration of anosmia, and recovery rates (01).
In a pilot study (neither randomized nor blinded), persistently dysosmic patients were treated with either a 10-day course of oral corticosteroids combined with olfactory training or with olfactory training alone (137). Nine participants received oral corticosteroids and performed olfactory training, and 18 performed olfactory training only. Only participants in the first group significantly improved their olfactory score and did so above the minimal clinically important difference for subjective smell improvement. Three participants who received oral corticosteroids reported minimal and transient side effects. This study provides weak support for oral corticosteroids in treating COVID-19-related olfactory disorders.
A retrospective, hospital-based, observational study of 141 patients in Italy found that only oral corticosteroids predicted 3-month recovery from chemosensory dysfunction (143).
Although systemic corticosteroids are considered a therapeutic option (98), others have recommended a more cautious approach until the results of randomized controlled trials are available (102). Reasons for not using systemic corticosteroids for COVID-19-associated chemosensory dysfunction include the following: (1) evidence supporting their usefulness is weak; (2) the rate of spontaneous recovery of COVID-19-related dysosmia is high; and (3) corticosteroids have significant potential adverse effects (102).
Olfactory training. A systematic review and metaanalysis found that olfactory training, in both the acute and chronic phases, improved olfactory disorders caused by COVID-19 (109).
Pentasodium diethylenetriamine. Because calcium cations are necessary for feedback inhibition concerning odor transmission, reducing free intranasal calcium cations using topical chelators might lead to restoration of the olfactory function in patients with post-COVID-19 anosmia. Consequently, a randomized controlled trial was conducted to test this idea: the topical chelator pentasodium diethylenetriamine pentaacetate (DTPA) significantly improved measured olfactory function compared to the control group in subjects with dysosmia ranging from functional anosmia to hyposmia post COVID-19 (111).
Gabapentin. In a randomized clinical trial, gabapentin was not associated with statistically significant or clinically meaningful benefit over placebo and likely is not an efficacious therapy for COVID-19-induced olfactory dysfunction (146).
Dysosmia in alpha-synucleinopathies. Olfaction in patients with Parkinson disease does not improve with levodopa therapy (105; 199), apparently at least in part because dopamine inhibits olfactory transmission in the olfactory glomeruli, and there is a paradoxical increase in the number of periglomerular dopaminergic neurons in the olfactory bulb in Parkinson disease (105).
All contributors' financial relationships have been reviewed and mitigated to ensure that this and every other article is free from commercial bias.

Douglas J Lanska MD MS MSPH
Dr. Lanska of the University of Wisconsin School of Medicine and Public Health and the Medical College of Wisconsin has no relevant financial relationships to disclose.
See ProfileNearly 3,000 illustrations, including video clips of neurologic disorders.
Every article is reviewed by our esteemed Editorial Board for accuracy and currency.
Full spectrum of neurology in 1,200 comprehensive articles.
Listen to MedLink on the go with Audio versions of each article.
MedLink®, LLC
3525 Del Mar Heights Rd, Ste 304
San Diego, CA 92130-2122
Toll Free (U.S. + Canada): 800-452-2400
US Number: +1-619-640-4660
Support: service@medlink.com
Editor: editor@medlink.com
ISSN: 2831-9125
General Neurology
Jan. 23, 2025
General Neurology
Jan. 13, 2025
General Neurology
Jan. 13, 2025
General Neurology
Jan. 13, 2025
Neuro-Ophthalmology & Neuro-Otology
Jan. 08, 2025
Neuro-Ophthalmology & Neuro-Otology
Jan. 07, 2025
General Neurology
Dec. 30, 2024
General Neurology
Dec. 28, 2024