


Movement Disorders
Acquired hepatocerebral degeneration
Jan. 20, 2025
MedLink®, LLC
3525 Del Mar Heights Rd, Ste 304
San Diego, CA 92130-2122
Toll Free (U.S. + Canada): 800-452-2400
US Number: +1-619-640-4660
Support: service@medlink.com
Editor: editor@medlink.com
ISSN: 2831-9125
Toll Free (U.S. + Canada): 800-452-2400
US Number: +1-619-640-4660
Support: service@medlink.com
Editor: editor@medlink.com
ISSN: 2831-9125
Worddefinition
At vero eos et accusamus et iusto odio dignissimos ducimus qui blanditiis praesentium voluptatum deleniti atque corrupti quos dolores et quas.
Historically, imaging studies were used experimentally to expand our knowledge and understanding of the pathophysiology and genesis of movement disorders. More recently, a few of these imaging tools have become part of clinical practice. In this article, we will address some of the key findings across imaging modalities in Parkinson disease, essential tremor, Huntington disease, dystonia, and Tourette syndrome.
|
• Structural and functional imaging studies are increasing our understanding of the underlying pathophysiology of movement disorders and beginning to show some utility in their ability to aid diagnosis and inform treatment development. | |
|
• Neurochemical imaging can be used to visualize nigrostriatal dopaminergic denervation in Parkinson disease. It may allow for early diagnosis and monitoring of disease progression, whereas the structural and functional changes in Parkinson disease may enable the ability to differentiate between motor and cognitive subtypes and to distinguish the disease from atypical parkinsonian syndromes. | |
|
• Advanced imaging techniques can be used to characterize the altered structure and aberrant function present within tremor-related cerebellar pathways in essential tremor and may help elucidate the presence of different phenotypes of this common movement disorder. | |
|
• Various imaging modalities have shown promise in detection of structural, functional, and neurochemical changes in pre-manifest Huntington disease and could support the development of disease-modifying therapies for this devastating neurologic disease. | |
|
• Although application of a broad range of imaging approaches is expanding our knowledge of the structural and functional changes associated with dystonia, additional larger and better-controlled studies are needed to unravel the complex physiology leading to this disabling disorder. | |
|
• Signaling pathway changes and the disruptions in motor, limbic, and cognitive circuit function that underlie the tics and other neuropsychiatric symptoms of Tourette syndrome may be able to be elucidated using modern neuroimaging techniques. |
Advances in neuroimaging techniques, including MRI, PET, and SPECT, have greatly enhanced our ability to study the structural, functional, and neurochemical brain changes that accompany and cause movement disorders.
Structural analysis of the brain can be performed with high-resolution anatomical MRI (up to 7 Tesla), voxel-based morphometry, and diffusion tensor imaging (DTI). Anatomic MRI scans can be used to examine volumes and shapes of particular brain structures, whereas voxel-based morphometry applies a statistical analysis of MRI to estimate brain volume changes systematically. DTI depicts molecular motion, including the direction of such motion, with results expressed in terms of mean diffusivity and fractional anisotropy. Diffusion tensor imaging data can also be used to estimate white matter pathways and their integrity using tractography (116).
Functional analysis of the brain can be performed with scintigraphy imaging (PET/SPECT) and functional MRI (fMRI). These functional brain imaging techniques can measure regional cerebral blood flow, glucose metabolism, and blood-oxygen-level dependent (BOLD) changes in the brain. Functional brain imaging identifies sites of regional pathology but generally lacks neurochemical specificity. Analysis of regional brain functions can be made more specific by performing repeated imaging before and after specific pharmacological interventions or specialized motor or mental tasks (“brain activation studies”).
Neurochemical imaging studies, which can be performed with PET, SPECT, and magnetic resonance spectroscopy (MRS), represent additional means by which we can investigate the pathophysiology of movement disorders. PET and SPECT imaging provide the ability to study the neurochemical, hemodynamic, or metabolic processes underlying movement disorders in vivo. MRS allows for a noninvasive method to determine neurotransmitter level (glutamate, glutamine, GABA, etc.) in a specific region of interest (246). These neuroimaging techniques allow the quantitative assessment of regional cerebral distribution of many molecules, proteins, enzymes, neurotransmitters, and neurotransmitter receptors in vivo.
Imaging investigations into the pathophysiology underlying movement disorders are frequently carried out, keeping in mind the motor basal ganglia and cerebellar circuitry. As our recognition and knowledge of associated nonmotor systems in movement disorders has expanded, affective and cognitive brain circuits have increasingly been examined with modern neuroimaging techniques.
Structural imaging in Parkinson disease. MRI imaging is increasingly utilized to distinguish the early stages of Parkinson disease from atypical parkinsonian disorders; however, anatomical differences have not been consistently identified (151). Using susceptibility-weighted imaging or T2 sequences, absence of dorsolateral nigral hyperintensity can be visualized and may help improve diagnosis. The dorsolateral nigral hyperintensity is thought to correspond to nigrosomes-1, which are small clusters of dopaminergic cells within the substantia nigra that are prominently affected in Parkinson disease. In a meta-analysis review, the overall sensitivity and specificity of the absence of dorsolateral nigral hyperintensity for Parkinson disease versus controls was 97.7% and 94.6%, respectively (168), with no effect on the findings being related to aging (104).
Imaging the substantia nigra with 3T susceptibility-weighted MRI may reveal the typical “swallow tail” sign, which has been found to have relatively high specificity and sensitivity for nigral degeneration in Parkinson disease (239) and may distinguish Parkinson disease from essential tremor (208) and vascular parkinsonism (284). 7 Tesla iron sensitive T2* MRI imaging demonstrates dorsolateral substantia nigra hyperintensity in controls. This "swallow tail" region of hyperintensity is absent in Parkinson disease patients (169).
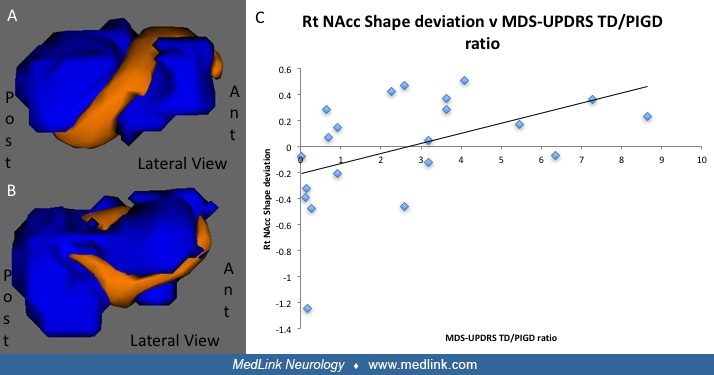
MRI can be used to evaluate changes in grey matter volume and shape that may aid in the diagnosis of Parkinson disease or be used to track progressive clinical features. For example, in one study of 21 patients and 20 healthy controls, shape deviations in the nucleus accumbens among the Parkinson disease patients compared to controls was driven by the tremor-dominant patients, raising the possibility that morphometric techniques may even be able to provide a means to distinguish between major motor subtypes (199). In cross-sectional studies evaluating cortical thinning in non-demented Parkinson disease patients, more severe disease was associated with progressive cortical thinning and subcortical volume loss (96; 272). Global cognitive performance also correlated with cortical thinning in frontal, temporal, parietal, and cingulate cortices as well as with hippocampal volume loss (272), suggesting that volumetric changes may be detectible in Parkinson disease prior to the onset of cognitive impairment.
DTI techniques have also been applied to evaluate the microstructural and white matter tract changes underlying Parkinson disease pathophysiology. Measuring microstructure alterations with DTI is potentially a valuable tool to use as a biomarker to aid in the diagnosis of Parkinson disease. Decreases in fractional anisotropy have been reported in the substantia nigra of patients with Parkinson disease (259; 283; 186) as well as in other subcortical and cortical regions (282; 283). A longitudinal study over 19 months demonstrated that nigral fractional anisotropy declines with disease progression, suggesting it may serve as an imaging measure of disease progression (156). Using DTI to assess whole-brain white matter structural connectivity has also shown promise in identifying structural changes to network topology that are present early on in the disease course of Parkinson disease (180).
Differences in DTI measures have also demonstrated some ability to distinguish Parkinson disease from atypical parkinsonian syndromes like multiple system atrophy and progressive supranuclear palsy (277; 130) and might be used to distinguish between Parkinson disease motor subtypes (186) and between cognitively intact and cognitively impaired patients with Parkinson disease (285).
A computational approach to DTI using a bi-tensor analysis model that allows for the calculation of the fractional volume of free water within a voxel may have application in neurodegenerative disorders (265). In using this imaging technique, free water was shown to increase over four years in the posterior substantia nigra of Parkinson disease patients, suggesting that the imaging approach may have utility in tracking disease progression (42). A separate group investigating the technique in a longitudinal study of 19 Parkinson disease patients and 19 healthy controls over three years found that free water within the anterior substantia nigra significantly increased over time (109). This method was also demonstrated in an international multicenter study of 1002 patients to distinguish those with Parkinson disease from atypical parkinsonian disorders with good accuracy using a machine learning-based approach (12). Results from these studies suggest that free water increases within the substantia nigra; this may be useful to monitor the progression of Parkinson disease as well as for clinical trials employing potentially disease-modifying therapies.
Retinal imaging is a relatively new modality that, in the future, may allow for early diagnosis as changes may precede the development of clinical signs of Parkinson disease. Challenges include a lack of large-scale clinical trials and variable sensitivity and specificity results among published trials. Spectral-domain optical coherence tomography can demonstrate thinning of the retinal nerve fiber layer and ganglion cell layer early in the disease (67).
Imaging in atypical parkinsonism.
Progressive supranuclear palsy. Midbrain atrophy on MRI imaging in progressive supranuclear palsy is an imaging marker most beneficial for active clinicians. A study of 23 progressive supranuclear palsy-Richardson syndrome (PSP-RS) patients and 42 Parkinson disease patients found 99% diagnostic accuracy when differentiating these disorders via midbrain area (280). Longitudinally, this marker is more accurate than 18F-AV-1451 (flortaucipir) PET ligand for measuring disease progression in clinical trials (268).
Corticobasal degeneration. Corticobasal degeneration (CBD) is manifested by frontal and parietal atrophy on MRI brain, usually contralateral to the more affected limb (236). 18F-flortaucipir has increased uptake in globus pallidus, premotor cortex, and motor cortex in corticobasal degeneration. There may also be off-target binding in the midbrain, possibly due to melanin-containing tissues (100).
Clinical diagnosis of corticobasal syndrome may be due to multiple possible pathologies, not just corticobasal degeneration but also progressive supranuclear palsy and others. A study of amyloid and tau binding in corticobasal syndrome patients showed the following results: amyloid positive and tau positive (Alzheimer-like) 13%, amyloid negative tau positive (69%), and amyloid negative and tau negative (19%). C-pyridinyl-butadienyl-benzothiazole 3 (11C-PBB3) was used for tau visualization, and C-Pittsburgh compound-B (11C-PiB) PET for amyloid visualization in this study (187).
Functional imaging in Parkinson disease. Principal component analysis of resting glucose metabolic studies has revealed the presence of a consistent topographic profile in Parkinson disease characterized by increased metabolic activity in the lentiform nucleus associated with decreased activity in the lateral frontal and premotor areas (73). Applying similar methodologies, a tremor-related pattern in Parkinson disease has been described and consists of increased activity within the thalamus, pons, premotor cortical regions, and putamen (11). A topographic profile using principal component analysis has been associated with executive functioning in nondemented patients with Parkinson disease and characterized by reduced metabolic activity in the pre-supplementary motor area, prefrontal cortex, precuneus, and parietal association regions and increased metabolic activity in the cerebellum (123). These functional imaging studies provide some insights into the abnormal networks that underlie specific clinical manifestations of the disorder and could provide a means to objectively measure responses to antiparkinsonian medications and interventions such as deep brain stimulation (15).
Functional connectivity analysis of resting state functional MRI (rsfMRI) data is an fMRI-based imaging technique with high spatial resolution that has also been increasingly applied to study network dysfunction in Parkinson disease. This neuroimaging technique can be used to identify brain regions exhibiting correlated patterns of spontaneous low-frequency oscillatory activity when participants are at rest and form consistent resting-state networks across participants (87). Using rsfMRI, changes to functional connectivity within corticostriatal networks have been examined in Parkinson disease and revealed that patients with Parkinson disease tend to have increased functional connectivity within some corticostriatal networks and decreased connectivity within other networks (278; 112). A metaanalysis of studies evaluating regional homogeneity and whole-brain rsfMRI used to examine local synchronization of spontaneous blood oxygenation-level-dependent signals revealed increased synchronization in the bilateral inferior parietal lobules, bilateral medial prefrontal cortices, and left cerebellum (202). Additionally, decreased synchronization was seen in the right putamen, right precentral gyrus, and left lingual gyrus. The regions of decreased synchronization are proposed to indicate a disease-related functional deprivation, whereas regions of increased synchronization may reflect a compensatory response (202). These findings are consistent with past rsfMRI studies that have shown increased connectivity in motor areas, including the cerebellum (243), as well as decreased activity in the right posterior putamen in Parkinson disease patients during motor tasks (115).
RsfMRI studies in Parkinson disease may be best suited, however, to investigate network disruptions associated with nonmotor symptoms such as cognitive impairment. In a metaanalysis that included 17 rsfMRI studies, for example, Parkinson disease patients with cognitive impairment showed a predominant pattern of reduced connectivity within key regions of the default mode network (274). This reduced connectivity may serve as a biomarker for cognitive impairment in Parkinson disease. RsfMRI has also been used to investigate psychiatric symptoms in Parkinson disease. For example, in a study of depression, it was found that Parkinson disease patients with mild-to-moderate depression had decreased regional homogeneity in the anterior cingulate cortex, and those with severe depression had increased regional homogeneity in the inferior prefrontal gyrus, orbitofrontal cortex, and supplementary motor area (264). Using an approach that evaluates dynamic functional connectivity within rsfMRI data, a study evaluating Parkinson disease patients with impulse control disorder found that these patients have diminished between-network connections but strong within-network connections in the salience network that correlated with the severity of impulsivity (188). These studies suggest that rsfMRI may be able to provide new insights into the distinct changes in brain function that contribute to psychiatric symptoms in Parkinson disease.
Graph theory analysis can be applied to rsfMRI data to investigate disruptions to more dispersed, large-scale functional brain networks in Parkinson disease. In one study, 21 patients with Parkinson disease were found to have widespread reduced functional connectivity but increased local efficiency, which reflects the network’s capacity for regional specialization within executive and salience resting state networks compared to 20 healthy controls (26). Dopaminergic therapy was also found to cause a significant reduction in whole brain local efficiency except for within the subcortical network, where it induced an increase in local efficiency. Even in early-stage patients who are drug-naïve, global and local functional integration and segregation has been found to be decreased in patients, and measures of network topology were found to correlate with clinical features, including motor severity and disease duration (245). In a separate study of 147 drug-naïve patients, functional brain network connectivity changes have further been found to predict cognitive progression over two years (65). Taken together, these findings suggest graph theoretical analyses of rsfMRI data can enhance our understanding of how whole-brain network functional organization is impacted by Parkinson disease and may provide a tool by which to track disease progression and responses to novel treatments under investigation.
Neurochemical imaging in Parkinson disease. Neurochemical imaging assessment of striatal dopaminergic innervation using PET or SPECT techniques in vivo has expanded our insight into the pathophysiology of Parkinson disease. Furthermore, these techniques have the potential to provide an objective and reliable measurement of nigrostriatal pathology and serve as a robust marker to study disease progression and evaluate therapies aimed at the preservation or restoration of nigrostriatal neurons. Many nuclear medicine tracers (radioligands) have been developed that can assess the presynaptic and postsynaptic dopaminergic system and quantify neuronal degeneration in movement disorders such as Parkinson disease by tracking the activity of various enzymes, transporters, and receptors (Table 1) (242; 192; 40; 232; 249).
Target | 11C | 18F | 123I | 99mTc | |
Presynaptic | VMAT2 | C-DTBZ | F-DTBZ | ||
DAT | CFT | F-CFT | I-altropane | TRODAT-1 | |
DA synthesis | F-DOPA | ||||
Postsynaptic | D1 | C-NNC 112 | F-fallypride | ||
D2/D3 | C-raclopride | I-IBZM |
Presynaptic dopamine transporter (DaT) imaging, largely employed with SPECT, has shown high sensitivity and specificity in differentiating Parkinson disease patients from healthy controls and patients with essential tremor. The DaT SPECT ligand Ioflupane I-123 was approved by the United States Food and Drug Administration in January 2011 as a means to assist in the diagnosis of adult patients with parkinsonian symptoms. Although DaT imaging has been used extensively in Europe and its use is rapidly expanding in the United States, its diagnostic value has limitations, including an inability to distinguish between Parkinson disease and other disorders with degeneration of nigral neurons and reliance on qualitative rather than quantitative visual assessment (21). Prior DaT imaging studies in parkinsonian conditions have found the sensitivity to range from 0.52 to 1.0, the specificity from 0.7 and 1.0, the diagnostic accuracy from 0.81 and 0.93, and the inter-rater reliability from 0.49 to 1.0. (62). Some investigators have raised concern about the possible redundancy of this type of imaging tool in diagnosing and managing patients with parkinsonian disorders (63). Nevertheless, DaT imaging, along with other means of assessing dopaminergic denervation in Parkinson disease, can detect early changes in Parkinson disease within the posterior putamen contralateral to the clinically most affected side (35) and provide a means for objective quantification of nigrostriatal integrity.
Although the rate of decline in presynaptic DaT levels varies by method and radioligand used, dopamine imaging has demonstrated an ability to track disease progression in Parkinson disease (198; 119). Additionally, striatal DaT binding imaging changes have also shown good agreement with postmortem studies of substantia nigral cell count irrespective of the cause of neuronal loss (145), but when looked at in a larger cohort of patients with Parkinson disease, there was no correlation found (230). It is possible that the link between DaT binding and nigral neuron loss becomes less accurate after neuronal losses exceed 50%, thus making DaT use for progression monitoring only accurate in mild to moderate disease (249). Investigations of DaT imaging in newly diagnosed patients with Parkinson disease suggest that declines in DaT uptake may also be linked to cortical thinning in frontal and posterior-cortical brain regions and correlate with declines in cognitive function (233). In a study of 54 nondemented patients, striatal DaT uptake correlated with impaired performance on memory and attention/executive tasks (86). These studies suggest dopaminergic imaging in the early stages of Parkinson disease could have potential clinical and therapeutic implications beyond motor symptom progression.
Also relevant for the study of neuroprotective therapeutic agents, dopamine imaging has shown a potential to detect pre-motor Parkinson disease. For example, decreased striatal DaT uptake has been reported in relatives of patients with Parkinson disease thought to be at risk for developing the condition (171), and decreased tracer uptake in the putamen has been reported in asymptomatic twins of patients with Parkinson disease (43). In a DaT imaging study of patients with idiopathic REM sleep behavior disorder, a subgroup known to be at higher risk of developing synucleinopathies such as Parkinson disease, reduced striatal uptake was seen in about 40% of patients and, when performed in combination with transcranial sonography, predicted the development of a synucleinopathy by 2.5 years in 8 of 27 patients (126). In a subsequent study by the same investigators involving 87 patients with polysomnography-confirmed REM sleep behavior disorder, a DAT deficit was found at baseline in 58.6% of the patients; during a follow-up period up to 10 years, 28.7% of these patients had developed a synucleinopathy with a mean latency of 3.2±1.9 years from imaging (127). In a study that included 38 patients with idiopathic REM sleep behavior disorder, 36.8% developed a synucleinopathy over 4.7 years, and the risk of this occurring was significantly elevated among those with reduced DaT-specific binding ratios in the putamen (51). These findings support that DaT imaging can identify REM sleep behavior disorder patients at short-term risk for Parkinson disease or other synucleinopathy.
The correlation of PET and SPECT measures of presynaptic dopaminergic activity with clinical severity in Parkinson disease suggests these imaging techniques could be useful biomarkers for monitoring disease progression or evaluating the effect of restorative or neuroprotective therapies. Their use in clinical trials to date, however, has generated conundrums. One issue is the identification of parkinsonian subjects who have scans without evidence of dopaminergic denervation (SWEDDs). Radiotracer-based imaging of the presynaptic nigrostriatal system has been utilized to support the clinical diagnosis of parkinsonism. However, in a couple of large neuroprotective studies in Parkinson disease using presynaptic nigrostriatal imaging as an outcome measure, approximately 10% to 15% of subjects were found to have normal dopaminergic scans (269; 80). Controversy remains about whether these SWEDDs represent misdiagnoses, tremor-variant syndromes, or limitations of the imaging techniques (182). Results from a 2-year follow-up study in 150 consecutive cases with SWEDDs supported that these subjects had nondegenerative movement disorder diagnoses in nearly all cases (175). In a case series, several SWEDD subjects clinically had asymmetric resting tremor and reduced arm swing and were re-diagnosed as having dystonic tremor (238); in the large prospective PRECEPT (Parkinson Research Examination of CEP-1347 Trial) study, the 11.4% of patients who had SWEDDs had their diagnosis changed to a disorder other than Parkinson disease by the end of the study (172). Overall, longitudinal studies with SWEDDs to date support that subjects with normal dopamine imaging do not have a disorder associated with nigrostriatal degeneration.
To date, presynaptic neurochemical imaging has not demonstrated an ability to reliably differentiate Parkinson disease from atypical parkinsonian disorders. More recently, however, novel approaches such as intrastriatal gradient analyses have suggested that caudate nucleus-to-anterior putamen declines in striatal dopaminergic transmission detected using 18F-DOPA PET may be able to discriminate Parkinson disease from atypical parkinsonian disorders (248).
Nondopaminergic neurochemical imaging in Parkinson disease. A growing area of PET and SPECT imaging in Parkinson disease involves applying nondopaminergic neurochemical imaging (see Table 2) (232; 249; 260). In a SPECT study evaluating nicotinic acetylcholine receptors using the SPECT ligand [123I]-5-I-A-85380, widespread significant decrease was found in subcortical and cortical regions in patients with Parkinson disease without dementia (90). Reduced thalamic and cortical acetylcholinesterase activity has also been demonstrated in nondemented patients with Parkinson disease, and the reduction was more prominent in those patients with a history of falls (38). In a study of 94 patients with Parkinson disease using the vesicular acetylcholine transporter PET ligand [18 F]FEOBV, vesicular acetylcholine transporter expression was significantly reduced in the thalamus of those patients with falls and in the striatum and limbic regions in those patients with gait freezing (36). These findings raise the possibility that different cholinergic systems contribute to the pathophysiology of falls and freezing of gait in Parkinson disease.
Changes to the serotoninergic system in Parkinson disease have also been investigated using PET and SPECT. Reduced binding to 5-HT1A receptors in the raphe nucleus that correlated with tremor scores has been reported in Parkinson disease in one study using the radioligand [11C]WAY 100635, a selective antagonist for 5-HT1A (69). Similarly, a study found reductions in serotonin transporter binding in the striatum and raphe nucleus that correlated with postural and action tremor severity using the serotonin transporter ligand [11C]DASB (157). PET and SPECT imaging are further helping illuminate a role for serotonergic mechanisms in levodopa-induced dyskinesias in Parkinson disease. For example, a combined D2 dopamine receptor ([11C]raclopride) and serotonin transporter ([11C]DASB) PET imaging study found that striatal serotonergic terminals contributed to levodopa-induced dyskinesias in Parkinson disease via aberrant processing of exogenous levodopa and release of dopamine as a false neurotransmitter in the denervated striatum (216). Disruptions to the serotonergic system in early-stage Parkinson disease may even predict a higher risk of developing levodopa-induced dyskinesia (89).
Using [11C]DASB and principal component analysis, a Parkinson disease-related spatial covariance pattern has also been identified for the serotonergic system (88). This pattern was comprised of decreases in binding within the striatum and substantia nigra and correlated with DTBZ binding and disease duration. A consistent topographic profile of the serotoninergic system analogous to prior profiles demonstrated with FDG PET in Parkinson disease could provide broader insight into the impact on neurochemical systems beyond the dopaminergic system in Parkinson disease.
The involvement of other neurotransmitter systems, such as those involving glutamatergic and adenosine 2A, has also been supported by PET imaging studies (03; 224). In serotonergic studies of nonmotor symptoms, patients with Parkinson disease with depressive symptoms had relatively increased serotonin transporter binding in the amygdala along with some other brain regions compared to patients without depressive complaints (215). Increased 5-HT2A receptor binding has also been reported in ventral visual pathways and bilateral dorsolateral prefrontal cortex in those patients with visual hallucinations (17). This latter finding helps support the treatment of psychosis in Parkinson disease with 5-HT2A receptor antagonists.
|
|
Target |
11C |
18F |
|
Cholinergic neuron |
AChE |
MP4A | |
|
VAChT |
FEOBV | ||
|
nAChR |
Nifene | ||
|
mAChR |
NMPB |
FP-TZTP | |
|
Serotonergic neuron |
SERT |
DASB | |
|
5-HT2AR |
RWAY |
MPPF | |
|
5-HT1AR |
MDL |
Setoperon | |
|
Intracellular |
Tau |
C-PBB3 |
F-AV-1451 |
|
Amyloid |
C-PIB |
F-florbetaben |
Imaging of fibrillary beta-amyloid plaques has been performed in patients with Parkinson disease at risk of dementia using the radioligand [11C]PIB. In one study, even mild increases of cortical amyloid binding (levels still well below those seen in Alzheimer disease) had significant correlations with cognitive impairment in patients with Parkinson disease (211). In a study of 98 newly diagnosed drug-naïve patients with Parkinson disease who underwent [18F]florbetaben imaging, the presence of amyloid pathology was associated with a higher rate (16 of the 18 amyloid-positive) of mild cognitive impairment (201). Increased cortical amyloid binding has also been found to correlate with the severity of postural instability and gait difficulty in Parkinson disease (184). These findings could help explain why patients with the postural instability and gait difficulty motor phenotype of Parkinson disease are at greater risk for the development of dementia.
Progressive supranuclear palsy and corticobasal degeneration belong to a group of neurodegenerative disorders called tauopathies, where tau becomes aberrantly hyperphosphorylated and dissociates from microtubules, resulting in a progressive accumulation of intracellular tau aggregates (185). Several radioligands designed to target tau have been developed with more newly developed [18F]THK-5351 showing potential to be used to detect tau in regions specific to progressive supranuclear palsy, whereas the previously promising radioligand [18F]-AV-1451 has been found to be better at identifying the paired helical filaments in Alzheimer disease than the straight conformation in progressive supranuclear palsy (56; 128). Further research is needed to develop a consistent radioligand that is specific to progressive supranuclear palsy and corticobasal degeneration without off-target binding.
In studying synucleinopathies like Parkinson disease, multiple system atrophy, and dementia with Lewy bodies, it would be ideal if we could directly image aggregated alpha-synuclein. This has so far proved to be a challenging task in Parkinson disease for several reasons: there are multiple forms of alpha-synuclein aggregates, the toxic alpha-synuclein aggregates are thought to primarily exist in oligomeric form as opposed to fibrillar form, the peptides or antibodies used must effectively cross the blood-brain barrier and reach into the cells where most of the alpha-synuclein pathology is present, and the peptides or antibodies used must be generated using radioisotopes with sufficiently long half-lives to make suitable brain imaging agents (40). Nevertheless, eight patients with probable multiple system atrophy were studied using [11C]BF227 PET in one study, and increased activity was measured in the basal ganglia and white matter of patients compared to healthy controls (140). The sensitivity and specificity of this study suggests, however, that this imaging technique is not sufficient to diagnose multiple system atrophy. Additional research is still needed to develop useful and practical ways to image alpha-synuclein aggregates, but the identification of novel ligands that bind alpha-synuclein fibrils does not appear to be that far away (177).
Essential tremor is a very common movement disorder with a prevalence above 4% in adults over the age of 65 years (162). It is defined as an isolated action tremor syndrome of the bilateral upper limbs of at least three years’ duration that can include tremor in other locations (head, voice, or lower limbs) but is not prominently accompanied by other neurologic signs such as dystonia, ataxia, or parkinsonism (27). Postural tremor is often considered to be a secondary tremor and is typically of lower amplitude than the kinetic tremor (161). Essential tremor is slowly progressive, with worsening and often debilitating tremor, though time course and severity are variable. The disease itself is heterogeneous, with variable development of parkinsonism, ataxia, and dystonia, which can make late-stage diagnosis difficult. This may result from multiple distinct phenotypes or distinct diseases, all characterized as essential tremor (66). At present, essential tremor remains a clinical diagnosis. Although essential tremor is commonly familial, no clear causative genes have been found, and the exact etiology of essential tremor remains unknown. Postmortem studies have been inconsistent, with some evidence of cerebellar neurodegeneration (55) and Lewy bodies in the brainstem (164). Whether pathologic variations are directly linked with phenotypic variations has yet to be definitively demonstrated. Imaging and physiological studies to date have implicated GABAergic abnormalities and dysfunction within the cerebello-thalamo-cortical network, often referred to as a “tremor network.” Biological markers and diagnostic tools are needed to improve our understanding of the cause of essential tremor, to aid in its diagnosis, and to promote the development of better treatments.
Structural imaging in essential tremor. Early voxel-based morphometry studies showed predominantly cerebellar and, to a lesser extent, cortical volume loss among patients with essential tremor. Subsequent studies showed diffuse cortical atrophy involving the posterior insular, superior temporal, and cingulate cortices (44). A metaanalysis from 2018 showed consistent reduction in the left precuneus extending to the left posterior cingulate gyrus (111). A metaanalysis of voxel-based morphometry studies in gray matter structures revealed heterogeneous results, leaving no evidence of changes in regional bray matter structures in essential tremor (165). Interestingly, a study evaluating young onset (before 30 years of age) essential tremor patients found increased cerebellar gray matter using voxel-based morphometry (222). The same group reported nearly identical findings in a larger group of patients younger than 60 years (45). Undoubtedly, heterogeneity of the essential tremor population and study methodology have made interpreting voxel-based morphometry studies quite difficult.
In a DTI study comparing 25 patients with familial essential tremor, 15 patients with Parkinson disease, and 15 controls, fractional anisotropy values in the dentate nucleus and the superior cerebellar peduncle were found to be significantly lower in the familial essential tremor group when compared with the other two groups (193). In a study with 57 essential tremor patients and 99 Parkinson disease patients, fractional anisotropy values were lower in the lateral geniculate body, striatum, pons, and internal capsule (134). Additionally, essential tremor patients were found to have greater diffusivity in the cerebellar peduncles. In another DTI study, essential tremor participants with (n=19) and without resting tremor (n=21) were found to have increased mean diffusivity of the cerebellar white matter amongst participants compared to 40 healthy controls (218). This finding suggests that white matter microstructure may be more altered in essential tremor with resting tremor and that this phenotype may be a distinct variant from essential tremor without resting tremor. In a small study with 12 essential tremor patients, fractional anisotropy in the ventral intermediate nucleus of the thalamus was compared before and after MR-guided focused ultrasound thalamotomy (266). The authors further identified a positive correlation between clinical response and reduction of fractional anisotropy after the procedure but fractional anisotropy before the surgery was not significantly different between those with and without clinical benefit.
Taken together, these findings support that DTI may have the ability to distinguish patients with essential tremor from those with Parkinson disease and may have some utility in advanced therapeutics after more in depth study.
Overall, structural imaging studies have largely implicated the cerebellum as the central pathological site in essential tremor, though more recent studies are beginning to reveal broader anatomical involvement. Whether the variability in results is because of multiple essential tremor phenotypes or noncomparable study design or patient variability remains unclear (240). This area of evaluation would benefit from larger studies both with a homogeneous essential tremor population (longstanding, familial, without parkinsonism, etc.) and robust subgroup analysis, including head tremor, voice tremor, and tremor severity.
Functional imaging in essential tremor. Early PET rCBF studies found increased perfusion of the cerebellum and supplementary motor area in essential tremor participants when manifesting tremor compared to controls (58). Subsequent studies identified additional areas with increased perfusion, including the thalamus, striatum, somatosensory cortex, and red nucleus (129; 270; 271).
BOLD fMRI studies in essential tremor have identified abnormal activity within the cerebello-thalamo-cortical network at rest (279; 195) and during the performance of motor and cognitive tasks (189). BOLD fMRI has shown abnormal changes that extend beyond the cerebello-thalamo-cortical network to the parietal cortex, primary visual cortex, and supplementary motor areas with visual feedback affecting the severity of tremor as well as the BOLD signal (13). In one study, reduced activation in the cerebellum, default mode network, and sensory-motor network was also found to correlate with tremor severity (92). Other BOLD studies using alternative motor tasks (eg, finger taps, figure 8s, etc.) have also showed reduced cerebellar activation in patients with essential tremor when compared with controls (41; 194; 92). One possible explanation for the presence of decreased cerebellar activity in essential tremor is that it represents a compensatory response to the elevated resting activity. Alternatively, reduced cerebellar activity during motor tasks could be a consequence of increased or supplementary motor activity.
In a multimodal MRI (voxel-based morphometry, DTI-tractography, rsfMRI) study of 19 patients with essential tremor and 19 controls, there was evidence of both structural and functional abnormalities in the cerebellum and supplementary motor area in patients with essential tremor (94). Relative to controls, patients with essential tremor had more supplementary motor area gray matter with less connectivity to the primary motor cortex. Structural and functional changes in the supplementary motor area correlated with clinical severity. The authors postulated that the increased gray matter and reduced connections in the supplementary motor area might stem from a compensatory attempt at directly modulating motor output via corticospinal projections instead of communicating directly with the motor strip.
A rsfMRI study of 25 essential tremor patients and 26 controls used seed analysis to investigate the functional connectivity of the dentate nucleus (279). Patients were found to have reduced functional connectivity between the dentate nucleus and structures in the frontal and parietal cortices bilaterally and subcortical grey matter (bilateral thalami and left caudate). Functional connectivity with the frontal and parietal cortices negatively correlated with severity of tremor and functional connectivity of subcortical structures negatively correlated with tremor frequency. This study echoes previous work that has identified the cerebellum playing a large role in tremor generation.
This collective body of research has identified abnormal cerebellar activity in the resting state of patients with essential tremor and more diffuse abnormalities, often associated with cerebellar projections when in an active state. Abnormalities in primary cortices, association cortices, and subcortical white matter structures have been identified.
Neurochemical imaging in essential tremor. A number of SPECT studies performed on patients with essential tremor have focused on DaT activity and utilized various radioligands. Although DaT imaging can be helpful in differentiating Parkinson disease from essential tremor, and most of these studies have not shown abnormalities in the basal ganglia, a few have found decreased uptake, indicating possible loss of dopaminergic neurons (50). For example, some studies found that patients with mixed tremors (large amplitude postural-kinetic tremors and low-amplitude resting tremors) have reduced striatal DaT binding (150; 68; 99). It is unclear if such findings stem from limitations including small sample size and ligand variability or potentially from improper diagnosis or disease heterogeneity. In a study of patients with essential tremor with and without parkinsonian features, however, careful quantitative analysis showed that patients with essential tremor did have slightly lower striatal binding ratios compared to healthy controls, especially in the caudate nucleus (263).
Flumazenil is a benzodiazepine receptor antagonist used to visualize the distribution of the GABA-A receptor in the brain. Its use as a radioligand, [11C]flumazenil, in a PET study including eight patients with essential tremor and 11 controls revealed increased receptor binding in the cerebellum, lateral premotor cortex, and ventrolateral thalamus (a region that receives cerebellar input and where thalamotomy has been attempted to alleviate tremor) (33). Another [11C]flumazenil study found a positive correlation between cerebellar GABA-A receptor availability and tremor severity (102).
MRS studies have consistently found no statistical difference in GABA levels of patients with essential tremor compared to controls when evaluating the bilateral cerebellar dentate nuclei and thalami (163; 255). One possible explanation for this negative finding is that increased terminal sprouting of Purkinje cells occurs in essential tremor leading to retained GABA levels despite an expected reduction in total Purkinje cell number. Conversely, another MRS study showed an increase in glutamate + glutamine (Glx) in the bilateral thalami compared to controls, with suggestion that this represents increased excitatory transmission in the thalamus, contributing to tremor (18). In a separate small study of 10 essential tremor patients, the GABA/Glx ratio in the cerebellum correlated with tremor severity, highlighting that it may be an imbalance of excitatory and inhibitory neurotransmission that underlies the severity of tremor (255).
In summary, there is limited evidence for the involvement of the dopaminergic system in essential tremor, but a growing number of neurochemical studies support the presence of an increased availability of GABA receptors, with further studies and clarification needed about what possible effect, if any, the overall GABA and glutamate concentrations in the dentate nucleus and thalamus have on essential tremor.
Huntington disease is an autosomal dominant monogenic neurodegenerative disease caused by an abnormally amplified number of CAG trinucleotide repeats in the huntingtin gene on the short arm of chromosome 4. Chorea is the most commonly recognized motor symptom in adult Huntington disease, but psychiatric (eg, anxiety, depression, disinhibition) and cognitive symptoms (eg, impaired planning, inattention, memory deficits) are also characteristic features. Pathologically, the abnormal huntingtin gene causes aggregates of the huntingtin protein and widespread neurodegeneration that particularly affects GABAergic medium spiny neurons in the striatum, though a link between these aggregates and the neurodegeneration has not been clearly established (228; 47). Although the diagnosis of Huntington disease is generally made through clinical assessment, family history, and genetic testing, imaging findings such as caudate atrophy can be seen early in disease and support the diagnosis. Imaging research pursuits in Huntington disease are often aimed at characterizing brain changes before the onset of clinical symptoms and finding ways to track disease severity so pre-symptomatic individuals can be identified and the efficacy of therapeutic interventions monitored.
Structural imaging in Huntington disease. It has been well demonstrated that significant atrophy occurs in the striatum of patients with Huntington disease. Indeed, striatal volume loss has been noted in patients with Huntington disease prior to motor symptom onset and is commonly used clinically to help support the diagnosis. The degree of striatal atrophy in Huntington disease has also been reported to correlate with age of onset, disease duration, and number of CAG repeats (207; 226). In a cross-sectional MRI study of 23 patients with Huntington disease with MRI scanning performed twice (10 months between scans), a greater rate of atrophy of the caudate nucleus, putamen, and total basal ganglia was seen in patients with earlier onset of clinical disease (16). Putamen atrophy has also been shown to be statistically different at time of conversion to manifest symptoms of Huntington disease compared to non-converters (61). Brain atrophy in Huntington disease is not limited to the striatum, however, as cortical and white matter atrophy have also been reported (227; 107; 60). Additionally, brain iron content has also been shown to increase before motor onset in the striatum, globus pallidus (52), and external capsule (132).
In a review of DTI studies in Huntington disease, fractional anisotropy was frequently reported to be increased in the caudate, putamen, and globus pallidus in patients with Huntington disease irrespective of their stage in the disease process, though this was not always a consistent finding (155). It was postulated that the increase in fractional anisotropy was caused by selective degeneration in the striatum, resulting in a more organized striatal structure. Others, however, have reported significant decreases in the fractional anisotropy in the cortical spinal tracts, corona radiata, corpus callosum, external capsule, thalamic radiations, superior and inferior longitudinal fasciculus, and inferior frontal-occipital fasciculus in Huntington disease patients compared to controls and premanifest patients (231). Only limited data exist on longitudinal microstructural changes in Huntington disease using DTI, but these data suggest that DTI is able to detect changes over time (231). Specifically, changes in fractional anisotropy of the corpus callosum were identified in a small cohort when reevaluated after seven months (253).
Functional imaging in Huntington disease. FDG PET studies in patients with Huntington disease have found decreased glucose utilization in the caudate nucleus and putamen; such findings may be present before striatal atrophy and clinical manifestations of disease (146; 178). In one FDG PET study, hypometabolism in the striatum of premotor patients with Huntington disease was correlated with conversion to clinically symptomatic Huntington disease within eight years (114). In a FDG PET study, hypometabolism in the frontotemporal region correlated with the presence and severity of apathy (234). In addition to the striatal findings, hypometabolism was identified in the mediotemporal lobes, and hypermetabolism was seen in the occipital cortex, thalamus, and cerebellum (82). These hypermetabolic areas may stem from compensatory mechanisms, but they have not been shown to track disease progression (83).
In a large rsfMRI study with 183 Huntington disease gene mutation carriers and 78 healthy controls, functional connectivity within the putamen was found to negatively correlate with CAG repeat length and the connectivity between the putamen and the insula decreased with increasing CAG repeat length (77). In a separate rsfMRI study comparing participants with Huntington disease gene expansion (premotor) and others with a family history of Huntington disease but without gene expansion, those participants with gene expansion had hyperconnectivity of inputs from the striatum to the cerebellum and outputs of the cerebellum early in life that waned over time (256). This was in contrast to those participants without gene expansion who showed no change or a slight increase in connectivity over time. These findings are suggestive of neurodevelopmental abnormalities in addition to the neurodegenerative changes we typically associate with Huntington disease.
Neurochemical imaging in Huntington disease. As Huntington disease progresses, medium spiny neurons, GABAergic cells within the striatum, degenerate. Dopaminergic transmission is conducted by these neurons through D1 and D2 receptors, the majority of which are postsynaptic (160). Therefore, consensus has been that the postsynaptic dopaminergic system is more significantly impacted in Huntington disease than the presynaptic system. Postsynaptic dopaminergic dysfunction has been thoroughly investigated. PET studies have been performed in Huntington disease using ligands to identify activity in D1 and D2 receptors, including [11C]raclopride (D2 receptor density) and [11C]SCH-23390 (D1 receptor density); both have detected reduced receptor density in the striatum of patients with Huntington disease correlated with disease duration (101; 204). Additionally, a number of studies have shown a reduction of D1 receptors in the temporal cortex, associated with cognitive dysfunction (101; 204). Reduced D2 receptor binding has also been found in the amygdala, frontal cortex, and hypothalamus (101; 10; 204; 214). Many of these postsynaptic dopaminergic receptor reduction findings have also been detected in preclinical patients with Huntington disease (262; 261; 206).
In a [11C]dihydrotetrabenazine PET study of six akinetic-rigid patients with Huntington disease, 13 chorea-dominant patients with Huntington disease, and 64 controls, reduced uptake of a VMAT 2 binder was seen in the caudate and putamen in all patients with Huntington disease, though the reduction was more significant in the akinetic-rigid patients (37). Using [11C]beta-CIT PET to evaluate presynaptic dopaminergic reuptake, five patients with Huntington disease compared to five healthy controls had decreased uptake in the caudate (53%) and putamen (55%) (101). Furthermore, in a 2-year longitudinal [123I]FP-CIT SPECT analysis of four patients with Huntington disease, mean uptake decreased by 5.8% in the caudate and 9.6% in the putamen and tracked clinical decline (95). Although many of the PET and SPECT studies in Huntington disease have been small and not analyzed in a consistent manner, together the findings suggest pre- and postsynaptic dopaminergic signaling is impaired in Huntington disease and that aberrant dopaminergic signaling likely extends beyond the striatum.
Many imaging studies have also investigated the nondopaminergic neurochemical changes associated with Huntington disease. In one PET study of 13 preclinical patients with Huntington disease and 10 manifest patients, an inverse relationship between GABA-A receptor binding ([11C]flumazenil) and D2 receptor binding ([11C]raclopride) was found in the putamen of symptomatic patients with Huntington disease (148). This finding was interpreted as indicating that the decline in dopamine receptors in Huntington disease is accompanied by a compensatory upregulation of GABA receptors.
Evidence for a role of microglial activation in the pathogenesis of Huntington disease has been supported by a [11C]PK11195 PET study in 11 patients with Huntington disease, which found a significant increase in binding within the striatum that correlated with a decline in D2 receptor binding ([C11]raclopride) and disease severity (205). Increased microglial activation detected with [11C]PK11195 has also been reported in the striatum as well as cortical regions including the prefrontal cortex and anterior cingulate of preclinical patients with Huntington disease (254), suggesting that it could be an early pathogenic process in Huntington disease. Imaging of phosphodiesterase 10A (PDE10A), which is mainly expressed in striatal medium spiny neurons and regulates striatal signaling by hydrolyzing cyclic adenosine monophosphate (cAMP) and cyclic guanosine monophosphate (cGMP), has revealed loss of binding within the striatum of patients with Huntington disease (02). Reduced PDE10A has also been demonstrated in patients with pre-clinical Huntington disease (229). A role of neuroinflammation in Huntington disease has further been evaluated with measurement of translocator protein expression using [11C]PBR28 and simultaneous PET/MRI (158). This study revealed the presence of neuroinflammation in the putamen and pallidum of patients that correlated well with brain atrophy on MRI (158). Together, these findings suggest imaging markers for neuroinflammation could be used to inform the development of novel inflammation-targeting treatments for Huntington disease.
Dystonia is a movement disorder characterized by sustained or intermittent muscle contractions causing abnormal, often repetitive movements, postures, or both (04). Dystonic movements are often patterned, twisting, and initiated or worsened by voluntary action and can be tremulous. It can be classified by clinical characteristics such as age of onset, body distribution, temporal pattern (persistent vs. action specific vs. dopamine responsive), and associated features. Body distribution classification includes focal (blepharospasm, cervical dystonia, writer’s cramp, etc.), segmental (two or more contiguous body regions), multifocal (two noncontiguous or more body regions), hemidystonia (restricted to one body side), and generalized (trunk and at least two other body regions) (04). Dystonia can occur as an isolated disorder or be associated with other neurologic or systemic conditions. It can be sporadic, inherited, or acquired from many different causes, including certain medications and brain ischemia, injury, or infection. Early evidence identified the basal ganglia playing a key role in the pathophysiology of dystonia, but in a growing number of advanced imaging studies, abnormalities have been detected beyond the basal ganglia, including cortical regions such as the sensorimotor, parietal and cingulate cortices, the brainstem, thalamus, and the cerebellum. To account for the wide range of brain structures outside the basal ganglia that appear to lead to dystonic movements, dystonia may be best understood as a network disorder (131). Imaging research in dystonia has begun to help unravel the complex pathophysiology underlying this complex movement disorder, from its associated molecular abnormalities up to the level of large-scale network dysfunction. The review of imaging in dystonia here is focused primarily on those studies investigating idiopathic isolated dystonia and some inherited forms of isolated dystonia.
Structural imaging in dystonia. A variety of voxel-based morphometry studies have been performed on patients with idiopathic focal dystonia. Despite variability in the body region affected by dystonia (cervical, focal hand, blepharospasm), many studies have found a bilateral increase in putaminal gray matter compared to healthy controls (30; 70; 78; 220). Data regarding other brain regions with changes in gray matter volume, including the cerebellum, thalamus, brainstem, and other cortical areas, have been less consistent (71; 200; 251; 120; 103). Some of these discrepancies may be related to the phenotypic variability of dystonia (257). For example, cervical dystonia patients with head tremor are shown to have greater atrophy of the vermis than patients without head tremor (167). Additionally, changes in white matter microstructures medial to the pallidum have also been reported in association with dystonia severity and can show a trend toward normalization after botulinum toxin injections (32).
In a DTI study of 18 patients with cervical dystonia, 16 patients with blepharospasm, and 35 healthy controls, increased fiber coherence and more ordered tissue (increased fractional anisotropy) was seen in the putamen of the patients with cervical dystonia (79). In addition, this study showed less ordered tissue (increased mean diffusivity) in the prefrontal cortex, supplemental motor area, and corpus callosum in patients with cervical dystonia. As such, the basal ganglia findings might be attributed to increased cellular density and the changes in the corpus callosum reflective of a disruption of communication between the motor portions of the two hemispheres. In a DTI study with 20 cervical dystonia patients, 15 blepharospasm patients, and 30 controls, reduced fractional anisotropy was seen in the cerebellum and caudate of cervical dystonia patients compared to controls and blepharospasm patients, whereas reduced fractional anisotropy was seen in the globus pallidus internus and red nucleus of blepharospasm patients compared to cervical dystonia patients and controls (24). Additionally, using diffusivity measures, patients with cervical dystonia and blepharospasm could be distinguished from each other with high reliability. Similarly, network connectivity differences have been shown between blepharospasm and hemifacial spasms at baseline and after treatment with botulinum toxin using structural correlation network analysis. Results continued to support the importance of the sensorimotor cortex and the cerebellum within and specific to dystonia (54). Together, these studies suggest that microstructural changes within nodes of the basal ganglia and cerebellar networks may differ between distinct focal dystonia phenotypes.
Although there have been some consistent and overlapping findings reported in DTI studies of isolated focal dystonia to date, sometimes conflicting findings, along with the lack of any significant changes, have been reported (59; 39; 64; 120; 212). This inconsistency could stem from the lack of any detectable structural changes with our imaging methods or may be due to patient variability, low numbers of subjects, or differences in imaging acquisition and analysis methods (223).
Similar to Parkinson disease, standard anatomic brain imaging is generally normal in patients with isolated and idiopathic forms of dystonia. Acquired dystonia from such causes as brain injury, infection, ischemia, and drugs or toxins, however, are frequently associated with abnormal standard clinical MRI imaging. Likewise, inherited forms of dystonia can have characteristic findings on structural MRI. For example, in neurodegeneration with brain iron accumulation syndromes (NBIA), which include pantothenate kinase-associated neurodegeneration (PKAN), phospholipase-associated neurodegeneration (PLAN), fatty acid hydroxylase-associated neurodegeneration (FAHN), beta-propeller protein-associated neurodegeneration (BPAN), and neuroferritinopathy, patients frequently show evidence of iron deposition within the basal ganglia (106). Many of these syndromes even have distinct findings on standard T2-weighted, diffusion-weighted imaging or gradient-recalled echo MRI that can aid in making correct diagnoses (09).
In PKAN, a disease characterized by dystonia, weakness, and mental retardation, T2-weighted MRI classically shows hypointensity within the globus pallidi with a small central area of hyperintensity, often referred to as an “eye of the tiger” sign. Importantly though, this finding is not pathognomonic for PKAN as it can be seen in other neurodegeneration with brain iron accumulation syndromes and neurodegenerative disorders (149). In PLAN, which is associated with weakness that progresses to spastic quadriplegia, optic nerve atrophy, language regression, cerebellar optic chiasm atrophy, and globus pallidi hypointensity can be seen on MRI (106). FAHN, which has been associated with both leukodystrophies and hereditary spastic paraplegia, is often associated with hypointensity of the globus pallidi on T2-weighted MRI as well as thinning of the corpus callosum. BPAN, which was previously called static encephalopathy with neurodegeneration in adulthood (SENDA), clinically consists of global developmental delay in childhood, dystonia, parkinsonism, and impaired cognition (110). Early in the disease, iron deposition in BPAN is maximal in the substantia nigra and can be seen as hypointense on T2-weighted MRI, and later hypointensities can be seen in the globus pallidi on diffusion-weighted imaging and gradient-recalled echo imaging. T1-weighted imaging in BPAN shows a characteristic hypointensity in the substantia nigra flanked by horizontal lines of hyperintense signal. Neuroferritinopathy is an adult-onset NBIA and is characterized by dystonia, chorea, and cognitive decline (147). MRI in neuroferritinopathy can show a distinct bilateral cystic degeneration of the putamen and globus pallidus surrounded by a hypointense rim on T2-weighted imaging.
Although high-resolution T2-weighted MRI can greatly assist in diagnosing neurodegeneration with NBIA, it is important to note that hypointensity of the basal ganglia can be seen in a variety other neurologic disorders. In children, hypointensity of the basal ganglia may be seen in a variety of metabolic neurodegenerative disorders as well as following hypoxic-ischemic injury (288). In adults, hypodense basal ganglia have been associated with neurodegenerative disorders, including Parkinson disease, multiple system atrophy, Huntington disease, and multiple sclerosis, NBIA, and may also be seen in healthy older adults due to physiological age-related iron deposition in the basal ganglia (20). Most of the published MRI images in these disorders are based on 1.5T technology, but it is important to note that 3T or 7T MRI images may resemble these images even though the patients do not have any of these abnormal disorders.
Functional imaging in dystonia. In fMRI studies of patients with writer’s cramp performing dystonia-inducing tasks, increased activation of the basal ganglia, thalamus, sensorimotor cortex and cerebellum has been found in patients compared to controls (219; 122). In musician’s dystonia, increased activation of the bilateral putamen with rsfMRI has been reported, along with additional evidence of altered basal ganglia functional connectivity in relation to temporal precision of keystrokes (141). In patients with blepharospasm, there is increased activation of the putamen when spasms occur (237) as well as increased activation in the right postcentral gyrus and precuneus, left precentral gyrus, and left occipital cortex compared to controls during reflexive blinking (191). When compared to controls, patients with cervical dystonia had increased activation of the sensorimotor cortex and cerebellum during an isometric head rotation task (221), as well as reduced superior collicular activation (179). Interestingly, increased sensorimotor network activation has been shown to be decreased by the engagement of a sensory trick (235). Patients with spasmodic dysphonia were found to have increased activation of the basal ganglia, sensorimotor cortex, and cerebellum during a speech production task (142), though reduced functional connectivity of the left inferior parietal cortex, putamen, and bilateral premotor cortices has been reported even when not speaking of resting state fMRI (19).
Overall, resting state and task-based fMRI studies suggest that dystonia symptoms are associated with abnormal activation within the basal ganglia, cerebellum, and primary and secondary sensorimotor processing brain regions and some degree of phenotypic specific alterations (159; 29). Although network dysfunction is hypothesized to contribute to the pathophysiology of dystonia, an rsfMRI study that included varied types of isolated focal dystonia supports the functional connectivity alterations involving these sensorimotor processing regions may be regional rather than global (197). These observed alterations could potentially serve as an endophenotypic marker for dystonia development in unaffected relatives of dystonia patients. Indeed, in one study alterations in functional connectivity of prefrontal-parietal cortex, thalamus, and caudate nucleus were noted in patients with laryngeal dystonia as well as their unaffected relatives but not in unrelated healthy controls (139). It should be noted, though, that when performing fMRI, likely pathophysiologic disturbances within dystonia from alterations in inhibitory signaling and dopaminergic neurotransmission may impact neurovascular coupling and alter the interpretation of fMRI. Thus, the hemodynamic response within dystonia may need to be accounted for differently than for subjects without dystonia (23).
FDG PET studies in patients with cervical dystonia have found increased glucose metabolism in the basal ganglia, motor cortices, thalamus, and cerebellum compared to controls (91; 166). These were similar findings to those reported in patients with blepharospasm, with the addition of hypermetabolism seen in the pons (76; 124; 252). FDG PET has also been used to investigate abnormal brain activation patterns in patients with DYT1, DYT6, and idiopathic blepharospasm using principal components analysis to identify spatial covariance patterns (74; 72; 124). One metabolic network pattern described was characterized by increased metabolic activity in the putamen, globus pallidus, cerebellum, and supplementary motor area. This pattern was identified in DYT1 carriers irrespective of whether clinical manifestations were present or not. A second metabolic network pattern in DYT1 patients was characterized by increased metabolic activity in the midbrain, cerebellum, and thalamus (72). This latter pattern was present only in those patients manifesting dystonia symptoms. In a regional cerebral blood flow PET study in six DYT1 patients and six controls imaged at rest and while performing paced joystick movements, DYT1 patients showed significant overactivity in the contralateral premotor cortex, rostral supplementary motor area, anterior cingulate, ipsilateral dorsolateral prefrontal cortex, and bilateral lentiform nuclei when compared to controls (48). Significantly reduced activity was found in the caudal supplementary motor area, bilateral sensorimotor cortices, and posterior cingulate and mesial parietal cortices. There were similar findings among patients with writer’s cramp (49; 125). These results are consistent with inappropriate over-activity of striatofrontal projections and impaired activity in motor executive areas and may explain the simultaneous dystonic posturing and bradykinesia evident in this disorder.
An important caveat to interpreting findings from functional imaging studies in dystonia is the potential influence and confounding effect on results of the presence of dystonia and sensory system engagement. Furthermore, the effect of botulinum toxin injections on the sensorimotor system and their potential to alter functional imaging results must be considered. Indeed, in a motor task-based fMRI study of 12 botulinum toxin naïve cervical dystonia patients, a significant increase in activation of the bilateral primary and secondary somatosensory cortex, bilateral superior and inferior parietal lobule, ipsilateral thalamus, putamen, and cerebellum amongst other regions was detected four weeks following their first set of injections (190). Furthermore, it has been reported that a single, first time injection of botulinum toxin for treatment of cervical dystonia can result in a reduction in excessive functional connectivity between the sensorimotor cortex and right superior frontal gyrus after only four weeks (84). Nevertheless, taken together, functional imaging data to date support the presence of overactivity of the basal ganglia in dystonia as well as the involvement of more distributed sensorimotor and cerebello-thalamo-cortical brain networks.
Neurochemical imaging in dystonia. Disruptions to presynaptic and postsynaptic dopaminergic signaling in dystonia have been investigated using both SPECT and PET with a variety of radioligands. These molecular imaging studies have consistently found decreased striatal D2 receptor binding in cervical dystonia (118), writer’s cramp (121), and cranial dystonia (blepharospasm and oromandibular dystonia) (209), as well as inherited forms of dystonia (DYT1 and DYT6) (14; 46). Because most of the D2-like radioligands used for these studies also bind D3 receptors, it is not presently known if these findings could be influenced by a decrease in striatal D3 receptor binding (135). Fewer studies have been conducted using radioligands that bind D1 receptors. In a study using a selective D1-like receptor ligand, no changes in striatal binding were seen in a mixed group of patients with focal dystonia compared to healthy controls (136).
A potential confound in D2/D3 binding studies in dystonia could be the presence of an underlying defect in striatal dopamine neurotransmission. In a [11C]raclopride study of patients with writer’s cramp, striatal dopamine release was decreased during a hand motor task and increased during an unaffected speech task (22). In an analogous study of patients with laryngeal dystonia, dopamine release was decreased during a speech task and increased during an unaffected hand motor task (241). Furthermore, in patients with cervical dystonia, reductions in striatal DaT and D2/3 binding potential were seen to correlate with depression (287). Overall, these data suggest that dystonia is likely associated with dysfunction of the D2 receptor-mediated indirect pathway, leading to loss of inhibition of striato-thalamo-cortical projections and unhampered motor activation.
Although striatal dopaminergic dysfunction is believed to play a significant role in many dystonias, other neurotransmitters and signaling systems may be involved in the pathogenesis of dystonia. In a [123I]iodobenzovesamicol SPECT study examining cholinergic terminal density in patients with cervical dystonia, striatal uptake was reduced compared to controls, supporting that dystonia is associated with abnormal cholinergic interneuron function within the striatum (05). Serotonergic abnormalities have also been investigated in cervical dystonia using [11C]DASB SERT (244). Although no significant differences were seen on direct comparison of patients to controls, higher binding potential in the dorsal raphe nucleus was positively correlated with motor symptom severity, pain, and sleep disturbances (244). In a small double-blinded placebo-controlled [123I]FP-CIT SPECT study evaluating the effects of escitalopram on patients with cervical dystonia, six weeks of escitalopram use did not lead to any significant differences in serotonin transporter binding but patients with subjective improvement in dystonia or mood symptoms did show a trend toward higher extrastriatal binding (286).
In a [11C]flumazenil PET study investigating GABA-A receptor binding in a mixed cohort of patients with isolated dystonia, a reduction in binding was found in the primary and secondary sensorimotor cortex and in the motor component of the cingulate gyrus (97). In a more recent [11C]flumazenil PET study of 18 patients with focal hand dystonia and 18 healthy controls, reduced GABA-A binding was also detected in the sensorimotor cortex, along with reduced binding in the cerebellum, insula, and inferior frontal operculum, and increased binding in the inferior frontal gyrus (93). In a [11C]flumazenil PET study with 15 cervical dystonia patients and 15 controls, however, increased GABA-A binding was detected in the motor cortex and parahippocampal gyrus (25). Additionally, GABA-A binding in the cerebellum negatively correlated with dystonia severity. Taken together, these studies support that abnormal GABA signaling within motor cortical and cerebellar pathways likely plays a role in isolated dystonia but that the relationship may be complex.
Additional evidence of GABAergic dysfunction underlying dystonia was revealed in a GABA MRS study of focal hand dystonia that found reduced GABA concentration within the basal ganglia and sensorimotor cortex in patients compared to controls (154). This finding, however, was not duplicated when an MRI system with a stronger magnet was used (113). Although a third GABA MRS study also did not find reduced GABA levels in patients with upper limb dystonia compared to controls, a difference in how GABA concentration was modulated in the motor cortex following repetitive transcranial magnetic stimulation between the groups was seen (173). GABA+/Cre levels were found to be decreased in the thalamus of patients with cervical dystonia compared to controls (108). Thus, dystonia may be associated with alterations in GABAergic neurotransmission as well as differences in GABA concentration at rest, depending on its phenotypic manifestation.
Tourette syndrome is a complicated neuropsychiatric disorder characterized by the childhood onset of motor and phonic tics (57). Tic disorders are the most common movement disorder in children, found in approximately 1% of young people ages 5 to 18 years old (143). Both motor and phonic tics can be simple (eg, focal muscle contraction, eye blinking, limb or head jerking, throat clearing, and grunting) or complex (eg, gesturing, hopping, humming, and coprolalia). An individual’s tics may fluctuate in type, frequency, and distribution over time. Twin and familial studies have indicated that Tourette syndrome has a genetic basis, and evidence supports polygenic inheritance (203; 176). Psychiatric comorbidities are extremely common in Tourette syndrome. The prevalence of obsessive-compulsive behaviors has been estimated to be 50% (08). Obsessive-compulsive behaviors are similar to tics in that they also have premonitory urges. Clinically, tics have been responsive to dopamine antagonists, whereas obsessive-compulsive behaviors are generally treated with selective serotonin reuptake inhibitors. Therefore, both serotonergic and dopaminergic pathways have been implicated in Tourette syndrome.
Structural imaging in Tourette syndrome. Studies evaluating structural changes in Tourette syndrome have focused on the basal ganglia, cortex, and corpus callosum (213). Many of the studies performed have been small and are not uniform in the age or gender groups tested, resulting in significant variability in findings. An MRI study of 47 patients with Tourette syndrome and 55 controls, all between the ages of 8 and 12 years, showed no significant difference between basal ganglia volumes (85). A prospective longitudinal study of 43 patients with Tourette syndrome evaluated symptoms and performed a structural MRI before age 14 years (average age = 11.4 years) and then re-evaluated symptoms and performed a structural MRI after the age of 16 (average = 18.7 years) (31). This study demonstrated a significant inverse correlation between caudate volume and severity of tics and obsessive-compulsive disorder symptoms and suggests that caudate volume may predict clinical outcome in patients with Tourette syndrome. This has been further supported by work showing a concordance between genetic risk for development of Tourette syndrome through single nucleotide polymorphism analysis and decreased caudate, thalamus, and putamen volume (183). Age-related structural changes specific to Tourette syndrome were also supported by a large, prospective MRI study of 155 patients with Tourette syndrome and 131 controls (210). In this study, patients with Tourette syndrome had reduced inferior occipital volumes and increased parieto-occipital volumes that were significantly associated with tic severity. Additionally, volumes of the dorsal prefrontal cortex were found to be increased in children with Tourette syndrome, and the volumes further increased with age, possibly representing compensatory changes. These findings are in contrast to evidence from a more recent structural MRI study showing reduced cortical thickness in the postcentral gyrus, superiorparietal gyrus, rostral anterior cingulate cortex in the left hemisphere and frontal pole, lateral occipital gyrus, and inferior temporal gyrus in the right hemisphere of 60 Tourette syndrome patients and 52 healthy controls (144). Additionally, lower white matter volume deep to the orbital and medial prefrontal cortex, along with increased gray matter volume in the posterior thalamus and hypothalamus, were reported in a study of 103 Tourette syndrome patients compared to healthy age-matched children (105).
In a DTI study of 27 children with Tourette syndrome and 27 controls, significantly reduced fractional anisotropy and increased radial diffusivity were found in the cortico-striato-thalamic-cortical circuit and superficial frontal white matter regions (267). Additionally, this study found a correlation between lower fractional anisotropy values (and higher radial diffusivity values) and increased Tourette syndrome symptoms. Similarly, structural connectivity between the globus pallidus internus with the limbic network, associative networks, caudate, thalamus, and cerebellum positively correlated with improvement in tics following deep brain stimulation for Tourette syndrome (133). These data suggest that white matter dysfunction in Tourette syndrome may extend beyond basal ganglia circuits and that an adaptive process is present within the motor and sensory cortical regions.
Functional imaging in Tourette syndrome. SPECT studies of regional cerebral blood flow in Tourette syndrome have shown reduced flow to the striatum and temporal and frontal lobes (181). FDG PET studies to date have typically been small but have largely shown reduced glucose metabolism in the striatum and increased metabolism in the sensorimotor cortex of patients with Tourette syndrome (225). An FDG PET study in 12 patients with Tourette syndrome and 12 controls, during which patients with Tourette syndrome were told not to suppress tics, identified increased metabolism in the premotor cortex and cerebellum and reduced uptake in the striatum in the Tourette syndrome group compared with the controls (217). In a [15O]water PET study of nine patients with Tourette syndrome and nine control subjects, activation of the insula, thalamus, putamen, and cerebellum was associated with tic release when compared to a sleep baseline (152).
Various fMRI designs have been used to study Tourette syndrome and the neural activation patterns associated with tic generation and suppression. One fMRI study with 22 adults with Tourette syndrome reported that tic suppression was associated with decreased activity in the globus pallidus and putamen and increased activity in the right caudate (138). In a more recent fMRI study with 40 Tourette syndrome patients and 20 controls, the superior temporal gyrus and midcingulate cortex were activated during the inhibition of urges, and greater premonitory urge severity was associated with increased activity in the hippocampus and middle temporal cortex as well (28). In a separate fMRI study with 66 patients with Tourette syndrome and 70 controls evaluated while subjects performed the Stroop test (a cognitive task that is known to activate fronto-striatal pathways), the posterior cingulate and prefrontal cortices became less activated with age in controls, but not the Tourette syndrome group (174). The diminished activation of posterior cingulate and prefrontal areas may suggest that patients with Tourette syndrome have less reflexive selective attention and processing speed with aging than the control subjects and further supports an alteration in functional connectivity that varies with age (196).
In a small event-related fMRI study with 10 adult patients with Tourette syndrome, tics were identified using synchronized video and audio recordings so that the neural correlates of tics and premonitory urges could be investigated (34). Activations within areas including the anterior cingulate and insular cortex, supplementary motor area, and parietal operculum were found to predominantly be activated before tic onset, whereas activations within sensorimotor areas, including superior parietal lobule and cerebellum, were found to be activated at the time of tic performance. Furthermore, similar brain regions within the premotor-parietal-cerebellar network were noted to be altered in Tourette syndrome patients compared to controls when assessing agency of movement (281). The severity of impaired measurement of agency correlated with motor symptoms, supporting that Tourette syndrome is associated with altered activation of the agency brain network. However, no changes in real-time fMRI were found within the supplementary motor area in subjects who had improvement in tic severity after undergoing neurofeedback compared to subjects who underwent sham neurofeedback training and did see improved tic severity (250).
Together these findings suggest that limbic and paralimbic pathways appear to be involved in tic generation and the urges and sensations that precede tics, whereas increased activity within sensorimotor pathways, including the basal ganglia and cerebellum, are associated with the performance of tics.
Neurochemical imaging in Tourette syndrome. Despite antidopaminergic medications having a therapeutic benefit in treating tics, a number of postsynaptic receptor binding studies in Tourette syndrome have not found D2 receptor density differences between patients with Tourette syndrome and control subjects (98; 137). Other studies, however, have found abnormalities in dopaminergic signaling in Tourette syndrome. In one PET study with 20 patients with Tourette syndrome, a small subgroup (4) with vocal tics had increased D2 receptor expression that correlated with the severity of tics (276). In a [I-123]IBZM SPECT study of monozygotic twins with Tourette syndrome, D2 receptor binding in the caudate nucleus was significantly greater in the clinically more affected twin, suggesting that increased D2 receptors contribute to the disease phenotype (273).
Presynaptic dopaminergic/DaT studies in Tourette syndrome have similarly found variable and, at times, inconsistent findings, with some studies showing no difference between patients and controls (258; 07; 01) and others showing increased uptake in the basal ganglia and brainstem in patients with Tourette syndrome (170; 75; 06; 53). In a meta-analysis of eight DaT binding studies involving 111 Tourette syndrome patients and 93 healthy controls, increased striatal binding was seen in patients; however, the difference was not statistically significant when the data were corrected for age and other differences between cohorts (117). There is also evidence of extrastriatal dopaminergic dysfunction in Tourette syndrome. For example, a [C11]FLB PET study in eight patients with Tourette syndrome and eight controls demonstrated reduced availability of D2/D3 receptor activity in cortical and subcortical regions, including the cingulate gyrus, insula, and thalamus in Tourette syndrome patients (247). The same group showed that an amphetamine challenge induced increased dopamine release in cortical regions, including the anterior cingulate and medial frontal gyri, in the patients with Tourette syndrome compared with controls. Overall, dopaminergic neurochemical studies support that Tourette syndrome is likely associated with a dysfunctional cortico-striato-thalamic-cortical circuit. It is unclear if the inconsistent findings to date stem from variable acquisition and analysis techniques or whether they are a consequence of the heterogeneity of the syndrome.
In addition to the dopamine-specific studies, there have been other studies of neurochemical systems in Tourette syndrome. For instance, in a serotonin transporter PET study in 11 patients with Tourette syndrome and 10 controls, reduced binding was found in the midbrain, caudate, and putamen in patients with Tourette syndrome (275). In a [11C]flumazenil study of 11 patients with Tourette syndrome and 11 healthy controls, widespread changes in GABA-A receptor binding were found, including decreased binding in the ventral striatum, globus pallidus, thalamus, amygdala, and insula and increased binding in the substantia nigra, periaqueductal gray, posterior cingulate cortex, and cerebellum (153). In a magnetic resonance spectroscopy study comparing patients with Tourette syndrome and obsessive-compulsive disorder with controls, no significant metabolite difference in the thalamus or dorsal anterior cingulate cortex of patients with Tourette syndrome was found, but glutamine levels in the region did correlate negatively with tic severity scores (81). These findings suggest that abnormalities in both the serotonergic and dopaminergic subcortical-cortical circuits are present in Tourette syndrome and that other neuro-metabolite differences may be present and could contribute to tic severity.
Modern imaging techniques, including MRI, DTI, fMRI, PET, and SPECT, have been used extensively to investigate movement disorders. By elucidating the key structural and functional findings associated with the disease process of Parkinson disease, essential tremor, Huntington disease, dystonia, and Tourette syndrome, we have gained a better understanding of the pathophysiology underlying symptoms in these distinct neurologic conditions. Neurochemical imaging of DaT is now used clinically to help confirm the presence of a neurodegenerative parkinsonian disorder, and other structural and functional imaging approaches are increasing our knowledge of the biological underpinnings of the heterogeneous motor and nonmotor symptoms caused by Parkinson disease. In essential tremor, structural imaging supports the key role that cerebellar dysfunction plays in the disorder, and neurochemical and functional imaging are helping us further characterize the roles that the GABAergic system and aberrant cerebello-thalamo-cortical network function play in the generation of tremor. A variety of structural, functional, and neurochemical imaging studies are expanding our knowledge of their biological underpinnings of the neurotransmitter signaling abnormalities and circuit disturbances that lead to hyperkinetic movements in Huntington disease and dystonia. In Tourette syndrome, imaging studies are just beginning to shed light on the pathophysiological changes associated with this complex neuropsychiatric disorder.
By addressing the limitations of many prior imaging studies and including larger sample sizes, recruiting homogenous populations, employing well-controlled scanning paradigms, and using robust analysis methods, imaging researchers will be able to bring coherence to prior conflicting imaging findings in movement disorders and pave the way for imaging markers to be developed that will inform treatment development and eventually aid the discovery of cures for these disabling neurologic diseases.
All contributors' financial relationships have been reviewed and mitigated to ensure that this and every other article is free from commercial bias.

Robert Fekete MD
Dr. Fekete of New York Medical College received consultation fees from Acadia Pharmaceutical, Acorda, Adamas/Supernus Pharmaceuticals, Amneal/Impax, Kyowa Kirin, Lundbeck Inc., Neurocrine Inc., and Teva Pharmaceutical, Inc.
See ProfileNearly 3,000 illustrations, including video clips of neurologic disorders.
Every article is reviewed by our esteemed Editorial Board for accuracy and currency.
Full spectrum of neurology in 1,200 comprehensive articles.
Listen to MedLink on the go with Audio versions of each article.
MedLink®, LLC
3525 Del Mar Heights Rd, Ste 304
San Diego, CA 92130-2122
Toll Free (U.S. + Canada): 800-452-2400
US Number: +1-619-640-4660
Support: service@medlink.com
Editor: editor@medlink.com
ISSN: 2831-9125
Movement Disorders
Jan. 20, 2025
Movement Disorders
Dec. 29, 2024
Movement Disorders
Oct. 24, 2024
Neurogenetic Disorders
Oct. 23, 2024
Peripheral Neuropathies
Aug. 22, 2024
Movement Disorders
Aug. 22, 2024
Neurobehavioral & Cognitive Disorders
Aug. 16, 2024
General Neurology
Aug. 14, 2024